|
4.3.6 đíŻY(jiĘŽ)
ŻY(jiĘŽ)║¤Ăń╦űĐđż┐Ň▀Á─Ë^Řc(diĘún)║═╬ĎéâÁ─îŹ(shʬ)˛×(yĘĄn)ŻY(jiĘŽ)╣ű���úČżC║¤┐╝Ĺ]│÷├š┬╩����íó│÷├šĽr(shʬ)Úg�����íó│÷├š╦┘┬╩íóĚÁăÓ┤Š╗ţ║═╔˙╬´┴┐Á╚ÄÎéÇ(gĘĘ)ÍŞś╦(biĘío)�úČ╬ĎéâŇJ(rĘĘn)×ÚĎ░╔˙║═ďď┼Óв▓ŢÁ─▀m║¤▓ąĚN╔ţÂ╚×Ú1-2cmúČÎţ╔ţ▓╗Ď╦│Č▀^(guĘ░)4cm�����íú▀@┼cĎďă░Đđż┐╦¨ŻĘÎhÁ─в▓Ţ▀m║¤▓ąĚN╔ţÂ╚4-5cm▓╗Ď╗Í┬����íúĎ░╔˙в▓ŢÁ─┐╣║ÁđďĆŐ(qiĘóng)Ë┌ďď┼Óв▓ŢúČĚÁăÓ┤Š╗ţ║═╔˙╬´┴┐?jĘę)?yĘşu)Ë┌ďď┼Óв▓Ţ�íúϲ┤╦úČď┌▀M(jĘČn)đđÍ▓▒╗╗ÍĆ═(fĘ┤)╣▄└ÝîŹ(shʬ)█`Ľr(shʬ)�����úČ▀xô˝║¤▀mÁ─▓ŢĚN║═▀m«ö(dĘíng)?shĘ┤)─▓ąĚN╔ţÂ╚╩ăĚă│ú▒ěϬÁ─íú
4.4 ┐Ţ▓ŢîËîŽ(duĘČ)в▓Ţ║═čo(wĘ▓)├ó╚ŞűťĚN├š│÷├š║═ďšĂ┌╔˙ÚL(zhĘúng)Á─Ë░ÝĹ
ďç˛×(yĘĄn)Ë┌2006─ŕ5-10ď┬ď┌║Ë▒▒╩íĆł╝Ď┐┌╩đ╚ű▒▒╣▄└Ýů^(qĘ▒)Íđç°(guĘ«)Ůr(nĘ«ng)śI(yĘĘ)┤ˇîW(xuĘŽ)▓ŢÁěĐđż┐îŹ(shʬ)˛×(yĘĄn)Ňżí¬í¬ç°(guĘ«)╝Ď▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ď░═Ô┐ĂîW(xuĘŽ)Đđż┐Ë^ťy(cĘĘ)ŇżúĘ╚ű▒▒úę▀M(jĘČn)đđ�����úČďç˛×(yĘĄn)ĚÍ×Ú╠´Úg║═ťě╩Ďâ╔▓┐ĚÍ�����íú╩Ď═Ôďç˛×(yĘĄn)ď┌╠ý╚╗▓Ţł÷(chĘúng)���úČ╩Ďâ╚(nĘĘi)ďç˛×(yĘĄn)ď┌îŹ(shʬ)˛×(yĘĄn)Ňżťě╩Ď▀M(jĘČn)đđ���íúâ╔ĚN┐Ţ▓ŢîË▓─┴¤ĚÍäe×ÚĆ─«ö(dĘíng)?shĘ┤)ě╠ý╚╗▓ŢÁě╩Ň╝»Á─┐Ţ┬ń╬´úĘ╠ý╚╗┐Ţ▓ŢúęĎď╝░╚╦╣Ą╦▄┴¤─úöM┐Ţ▓ŢîËúĘ─úöM┐Ţ▓ŢúęúČčo(wĘ▓)├ó╚ŞűťĚNÎË╝░Ď░╔˙в▓ŢĚNÎË×Ú2004─ŕď┌«ö(dĘíng)?shĘ┤)ě╠ý╚╗▓ŢÁě╩Ň╝»Á─ĚNÎË�����íúďç˛×(yĘĄn)ďO(shĘĘ)2ĚN▓╗═ČÁ─Ł▓╦«┴┐��íó 4ĚN▓╗═ČÁ─┐Ţ▓ŢîËÍě┴┐�����íó3ĚN▓ąĚN╬╗Í├║═2ĚN┐Ţ▓ŢîËţÉđ═íú
4.4.1 ┐Ţ▓ŢîËţÉđ═îŽ(duĘČ)ĚN├šŻĘÍ▓Á─Ë░ÝĹ
ťě╩ĎîŹ(shʬ)˛×(yĘĄn)▒Ý├¸úĘ▒Ý1-21║═▒Ý1-22úę�����úČ┐Ţ▓ŢîËţÉđ═îŽ(duĘČ)ĚN├š│÷├š║═╔˙ÚL(zhĘúng)╦┘Â╚Ëđ´@Í°Ë░ÝĹ�����íú╠ý╚╗┐Ţ▓Ţ┼c─úöM┐Ţ▓ޤÓ▒╚ŻÁÁ═┴╦в▓Ţ║═čo(wĘ▓)├ó╚ŞűťÁ─ĚN├š│÷├š┬╩║═╔˙ÚL(zhĘúng)╦┘Â╚���íú┤╦═Ô��úČ╠ý╚╗┐Ţ▓ŢĎ▓´@Í°ŻÁÁ═┴╦в▓ŢÁ─┤Š╗ţ┬╩║═Áě¤┬╔˙╬´┴┐����íú
4.2 Ł▓╦«┴┐îŽ(duĘČ)ĚN├šŻĘÍ▓Á─Ë░ÝĹ
Ł▓╦«┴┐îŽ(duĘČ)│÷├š┬╩���íó╔˙╬´┴┐║═╔˙ÚL(zhĘúng)╦┘Â╚Ëđ´@Í°Ë░ÝĹ����í����ú┐éÁ─üÝ(lĘói)Ív���úČŞ▀╣Ó╦«┴┐ď÷╝Ë┴╦čo(wĘ▓)├ó╚Şűť║═в▓ŢÁ─│÷├š┬╩║═╔˙╬´┴┐íúŁ▓╦«┴┐┤ˇÁ─ĚN├š│÷├š╦┘Â╚┐ýË┌Ł▓╦«┴┐đíÁ─úĘĎŐ▒Ý1-21║═▒Ý1-22úę��íú
4.3 ┐Ţ▓ŢîËÍě┴┐îŽ(duĘČ)ĚN├šŻĘÍ▓Á─Ë░ÝĹ
ťě╩Ďďç˛×(yĘĄn)▒Ý├¸úĘ▒Ý1-21║═▒Ý1-22úę����úČ┐Ţ▓ŢîËÁ─Íě┴┐´@Í°Ë░ÝĹв▓ŢÁ─╔˙ÚL(zhĘúng)╦┘Â╚║═Áě¤┬╔˙╬´┴┐úČ°îŽ(duĘČ)čo(wĘ▓)├ó╚ŞűťÁ─Ş¸ÝŚ(xiĘĄng)ÍŞś╦(biĘío)ż¨čo(wĘ▓)´@Í°Ë░ÝĹ��íúĚN├š╔˙ÚL(zhĘúng)╦┘Â╚║═┐é╔˙╬´┴┐ŰSÍ°┐Ţ▓ŢîËÍě┴┐Á─ď÷╝Ë°ď÷╝Ë�íúĎ░═ÔŚl╝■¤┬úĘ▒Ý1-23║═▒Ý1-24úęúČ┐Ţ▓ŢîËÍě┴┐îŽ(duĘČ)в▓ŢÁ─│÷├š┬╩║═┤Š╗ţ┬╩Â╝Ëđ´@Í°Ë░ÝĹ���úČîŽ(duĘČ)čo(wĘ▓)├ó╚ŞűťÁ─│÷├š┬╩║═┤Š╗ţ┬╩Ť](mĘŽi)Ëđ´@Í°Ë░ÝĹ���íúв▓ŢÁ─│÷├š┬╩ŰS┐Ţ▓ŢîËÍě┴┐Á─ď÷╝Ë°ŻÁÁ═úČÁź┤Š╗ţ┬╩ůsŰS┐Ţ▓ŢîËÍě┴┐Á─ď÷╝Ë°ď÷╝Ë�úČ┐╔─▄╩ăË╔Ë┌┐Ţ▓ŢîËËđ└űË┌▒ú│Í═┴╚└╦«ĚÍÁ─żë╣╩íú
4.4 ĚNÎË╬╗Í├îŽ(duĘČ)ĚN├šŻĘÍ▓Á─Ë░ÝĹ
▒Ý1-21���íó22║═23║═24´@╩ż�����úČĚNÎË╬╗Í├îŽ(duĘČ)ťě╩Ď║═Ď░═ÔÁ─в▓Ţ║═čo(wĘ▓)├ó╚ŞűťÁ─│÷├š┬╩Â╝Ëđ´@Í°Ë░ÝĹ����íú▓ąË┌═┴╚└ÍđÁ─ĚNÎË│÷├š┬╩▀h(yuĘún)▀h(yuĘún)Ş▀Ë┌Ăń╦Řâ╔ĚN▓ąĚNĚŻ╩ŻúČ°▓ąË┌┐Ţ▓ŢîË┼c═┴╚└▒Ý├ŠÍ«ÚgÁ─ĚNÎË│÷├š┬╩Ď▓┤ˇË┌▓ąË┌┐Ţ▓ŢîË▒Ý├ŠÁ─│÷├š┬╩���íúĚNÎË╬╗Í├îŽ(duĘČ)в▓ŢĚNÎË│÷├š║═╔˙ÚL(zhĘúng)Á─Ë░ÝĹ▒╚îŽ(duĘČ)čo(wĘ▓)├ó╚ŞűťÁ─Ë░ÝĹϬ┤ˇ�����íúĎ░═ÔŚl╝■¤┬Á─═┴╚└ÍđÁ─в▓Ţ│÷├š┬╩╩ăĚNÎË╬╗Ë┌┐Ţ▓ŢîË▒Ý├ŠÁ─2-3▒ÂÂÓ�úČčo(wĘ▓)├ó╚ŞűťÁ─│÷├š┬╩Í╗¤Ó▓ţ10%ΡËĎ���íúĚNÎË╬╗Í├îŽ(duĘČ)ťě╩Ďв▓Ţ║═čo(wĘ▓)├ó╚ŞűťÁ─╔˙ÚL(zhĘúng)╦┘Â╚����íóÁě╔¤���íóÁě¤┬║═┐é╔˙╬´┴┐Â╝Ëđ´@Í°Ë░ÝĹúČĚNÎË▓ąË┌═┴╚└ÍđÁ─öÁ(shĘ┤)ÍÁÎţŞ▀���íú
▒Ý1-21 ┐Ţ▓ŢîËţÉđ═��íóŁ▓╦«┴┐�����íó┐Ţ▓ŢîËÍě┴┐┼cĚNÎË╬╗Í├îŽ(duĘČ)čo(wĘ▓)├ó╚Şűť│÷├š║═┤Š╗ţÁ╚Á─Ë░ÝĹúĘťě╩Ďúę
|
╠Ä└Ý |
╦«ĂŻ |
│÷├š┬╩úĘ%úę |
┤Š╗ţ┬╩úĘ%úę |
╔˙ÚL(zhĘúng)╦┘Â╚úĘcmúę |
Áě╔¤╔˙╬´┴┐úĘgúę |
Áě¤┬╔˙╬´┴┐úĘgúę |
┐é╔˙╬´┴┐úĘgúę |
|
┐Ţ▓ŢîËţÉđ═ |
╠ý╚╗┐Ţ▓Ţ |
68.22b |
96.4a |
14.33b |
3.30a |
6.06a |
9.36a |
|
╦▄┴¤┐Ţ▓Ţ |
81.22a |
96.5a |
16.74a |
2.97a |
5.78a |
8.75a |
|
Ł▓╦«┴┐ |
Á═ |
72.67b |
96.08a |
16.46b |
2.94b |
6.05a |
8.99a |
|
Ş▀ |
76.78a |
96.76a |
14.61a |
3.34a |
5.79a |
9.13a |
|
┐Ţ▓ŢîË
Íě┴┐ |
0 |
75.72a |
96.67a |
15.36a |
2.93a |
5.56a |
8.49a |
|
1/2ĂŻż¨ÍÁ |
75.56a |
97.94a |
16.72a |
3.26a |
5.99a |
9.26a |
|
ĂŻż¨ÍÁ |
70.89a |
94.78a |
14.72a |
3.13a |
5.44a |
8.57a |
|
2▒ÂĂŻż¨ÍÁ |
76.72a |
96.31a |
15.35a |
3.23a |
6.68a |
9.91a |
|
ĚNÎË╬╗Í├ |
┐Ţ▓ŢîË▒Ý├Š |
60.88c |
98.31b |
14.36b |
2.63b |
4.79b |
7.42b |
|
┐Ţ▓ŢîË┼c═┴╚└Úg |
71.96b |
96.60ab |
14.68b |
2.90b |
5.05b |
7.95b |
|
═┴╚└Íđ |
91.33a |
94.35a |
17.56a |
3.88a |
7.91a |
11.80a |
Îóú║«ö(dĘíng)?shĘ┤)ě▓Ţďş┐Ţ▓ŢîËÍě┴┐ĂŻż¨ÍÁ×Úú?/SPAN>100g/m2�ú╗Ł▓╦«┴┐ĚÍäe×Ú«ö(dĘíng)?shĘ┤)ěŻÁËŕ┴┐ÎţđíÍÁ┼cÎţ┤ˇÍÁ╝┤50mm/ď┬║═100mm/ď┬
▒Ý1-22 ┐Ţ▓ŢîËţÉđ═íóŁ▓╦«┴┐���íó┐Ţ▓ŢîËÍě┴┐┼cĚNÎË╬╗Í├îŽ(duĘČ)Ď░╔˙в▓Ţ│÷├š║═┤Š╗ţÁ╚Á─Ë░ÝĹúĘťě╩Ďúę
|
╠Ä└Ý |
╦«ĂŻ |
│÷├š┬╩úĘ%úę |
┤Š╗ţ┬╩úĘ%úę |
╔˙ÚL(zhĘúng)╦┘Â╚úĘcmúę |
Áě╔¤╔˙╬´┴┐úĘgúę |
Áě¤┬╔˙╬´┴┐úĘgúę |
┐é╔˙╬´┴┐úĘgúę |
|
┐Ţ▓ŢîË
ţÉđ═ |
╠ý╚╗┐Ţ▓Ţ |
44.61b |
86.97b |
11.57b |
1.90a |
1.98b |
3.89a |
|
╦▄┴¤┐Ţ▓Ţ |
52.56a |
91.63a |
16.91a |
1.78a |
2.49a |
4.27a |
|
Ł▓╦«┴┐ |
Á═ |
38.78b |
89.67a |
12.64b |
1.61b |
2.08b |
3.69b |
|
Ş▀ |
58.39a |
88.93a |
15.84a |
2.08a |
2.39a |
4.47a |
|
┐Ţ▓ŢîË
Íě┴┐ |
0 |
50.39a |
76.14b |
12.41b |
1.66a |
1.74b |
3.40a |
|
1/2ĂŻż¨ÍÁ |
46.33a |
93.11a |
14.13ab |
1.74a |
2.18ab |
3.92a |
|
ĂŻż¨ÍÁ |
47.89a |
94.72a |
15.02ab |
1.91a |
2.56a |
4.47a |
|
2▒ÂĂŻż¨ÍÁ |
49.72a |
93.22a |
15.40a |
2.07a |
2.47ab |
4.53a |
|
ĚNÎË╬╗Í├ |
┐Ţ▓ŢîË
▒Ý├Š |
28.50c |
89.67a |
11.42b |
1.28b |
1.44b |
2.72b |
|
┐Ţ▓ŢîË
┼c═┴╚└Úg |
39.67b |
89.94a |
12.44b |
1.53b |
1.83b |
3.35b |
|
═┴╚└Íđ |
77.58a |
88.29a |
18.86a |
2.73a |
3.44a |
6.17a |
Îóú║«ö(dĘíng)?shĘ┤)ě╠ý╚╗▓ŢÁě┐Ţ▓ŢîËÍě┴┐ĂŻż¨ÍÁ×Úú?/SPAN>100g/m2�ú╗Ł▓╦«┴┐ĚÍäe×Ú«ö(dĘíng)?shĘ┤)ěŻÁËŕ┴┐ÎţđíÍÁ┼cÎţ┤ˇÍÁ╝┤50mm/ď┬║═100mm/ď┬
▒Ý1-23 ╠ý╚╗┐Ţ▓ŢîËÍě┴┐┼cĚNÎË╬╗Í├îŽ(duĘČ)čo(wĘ▓)├ó╚Şűť│÷├š║═┤Š╗ţÁ─Ë░ÝĹúĘ╠´Úgúę
|
╠Ä└Ý |
╦«ĂŻ |
│÷├š┬╩úĘ%úę |
┤Š╗ţ┬╩úĘ%úę |
|
┐Ţ▓ŢîËÍě┴┐ |
0 |
47.33a |
53.13a |
|
1/2ĂŻż¨ÍÁ |
51.47a |
50.67a |
|
ĂŻż¨ÍÁ |
53.47a |
47.27a |
|
2▒ÂĂŻż¨ÍÁ |
50.53a |
50.80a |
|
ĚNÎË╬╗Í├ |
┐Ţ▓ŢîË▒Ý├Š |
41.80b |
50.85a |
|
┐Ţ▓ŢîË┼c═┴╚└Úg |
41.20b |
52.15a |
|
═┴╚└Íđ |
50.70a |
48.40a |
Îóú║«ö(dĘíng)?shĘ┤)ě╠ý╚╗▓ŢÁě┐Ţ▓ŢîËÍě┴┐ĂŻż¨ÍÁ×Úú?/SPAN>100g/m2
▒Ý1-24 ╠ý╚╗┐Ţ▓ŢîËÍě┴┐┼cĚNÎË╬╗Í├îŽ(duĘČ)Ď░╔˙в▓Ţ│÷├š║═┤Š╗ţÁ─Ë░ÝĹúĘ╠´Úgúę
|
╠Ä└Ý |
╦«ĂŻ |
│÷├š┬╩úĘ%úę |
┤Š╗ţ┬╩úĘ%úę |
|
┐Ţ▓ŢîËÍě┴┐ |
0 |
30.80b |
18.87b |
|
1/2ĂŻż¨ÍÁ |
13.60a |
16.73b |
|
ĂŻż¨ÍÁ |
20.53ab |
31.47a |
|
2▒ÂĂŻż¨ÍÁ |
19.47ab |
33.40a |
|
ĚNÎË╬╗Í├ |
┐Ţ▓ŢîË▒Ý├Š |
12.90b |
25.10a |
|
┐Ţ▓ŢîË┼c═┴╚└Úg |
18.00b |
26.80a |
|
═┴╚└Íđ |
32.40a |
23.45a |
Îóú║«ö(dĘíng)?shĘ┤)ě╠ý╚╗▓ŢÁě┐Ţ▓ŢîËÍě┴┐ĂŻż¨ÍÁ×Úú?/SPAN>100g/m2
4.5 đíŻY(jiĘŽ)
1úę�íó▓╗═ČÁ─╣▄└Ý┤Ű╩ęúĘ░ĎÁěíóěÎŞţ���íó╩ęĚ╩�����íó▓ąĚN├▄Â╚úęîŽ(duĘČ)Đa(bĘ│)▓ą─▄Ě˝│╔╣ŽËđ´@Í°Á─Ë░ÝĹ����úČÍ▒ŻË╚÷▓ą���íóîŽ(duĘČ)Í▓▒╗▓╗▀M(jĘČn)đđö_äË(dĘ░ng)���úČ╝┤╩╣╩ęË├Ě╩┴¤║═ď÷╝Ë▓ąĚN├▄Â╚Ď▓║▄Űy╩╣Đa(bĘ│)▓ąÁ─╬´ĚNĚN├š│╔╣ŽŻĘÍ▓�����íú░ĎÁě?cĘói)_äË(dĘ░ng)┐╔Ďď═Ę▀^(guĘ░)ŻÁÁ═Í▓▒╗╔wÂ╚����íóď÷╝ËĚNÎË┼c═┴╚└ŻËË|��íóď÷ĆŐ(qiĘóng)┴╦╣ÔŇŇ║══┴╚└▒ÝîËťěÂ╚����úČĆ─°ď÷╝ËĚN├š│÷├š║═╔˙ÚL(zhĘúng)▒ÝČF(xiĘĄn)úČËđ└űË┌ĚN├š│╔╣ŽŻĘÍ▓�íú
2úęíóĚN├š├▄Â╚┼c▓ąĚN├▄Â╚│╩Ňř¤ÓŕP(guĘín)ŕP(guĘín)¤Á����úČĐa(bĘ│)▓ąĽr(shʬ)▀m«ö(dĘíng)╝Ë┤ˇ▓ąĚN├▄Â╚╩ă▒ěϬÁ─íú
3úę����íó▓╗═ČÁ─╬´ĚNĂńĐa(bĘ│)▓ąÂĘżË│╔╣ŽÁ─ÄÎ┬╩Ď▓▓╗═ČúČčo(wĘ▓)├ó╚ŞűťÁ─ĚN├šŻĘÍ▓─▄┴ŽĎ¬â×(yĘşu)Ë┌в▓Ţ�úČ▒ÝČF(xiĘĄn)×Ú│÷├š┐ýíó╔˙ÚL(zhĘúng)┐ý��úČ▓╗Ş▓═┴úĘ═┴╚└▓╗ö_äË(dĘ░ng)úę│÷├š┬╩Ď▓Ş▀Ë┌в▓ŢúČϲ┤╦čo(wĘ▓)├ó╚ŞűťĐa(bĘ│)▓ąŻĘÍ▓Á─đž╣űϬ║├Ë┌в▓Ţ��íú
4úę����íóĐa(bĘ│)▓ąĐ˛▓Ţ║═čo(wĘ▓)├ó╚ŞűťĽ■(huĘČ)╩▄ÁŻüÝ(lĘói)ÎďČF(xiĘĄn)┤ŠÍ▓▒╗Á─ĎÍÍĂθË├�����úČż▀ˇw▒ÝČF(xiĘĄn)×ÚĎÍÍĂ│÷├š║═¤ŮÍĂĚN├š╔˙ÚL(zhĘúng)�úČË╚Ăń╩ăüÝ(lĘói)ÎďČF(xiĘĄn)┤ŠÍ▓▒╗Á─Áě¤┬Şé(jĘČng)áÄ(zhĘąng)╩ă¤ŮÍĂĚN├šŻĘÍ▓Á─Îţ͸Ϭϲ╦ěíú
5úę��íó╗ý▓ą─┴▓ŢĂńĚN├šŻĘÍ▓▀^(guĘ░)│╠ÍđĚN├šÚgĎ▓Ëđ¤Ó╗ąÎ¸Ë├���úČčo(wĘ▓)├ó╚ŞűťĚN├š│÷├šďš��íó╔˙ÚL(zhĘúng)┐ý����úČîŽ(duĘČ)в▓ŢĚN├š╔˙ÚL(zhĘúng)ż▀ËđĎÍÍĂθË├íú
6úę��íó▓ąĚN╔ţÂ╚║═ĚNÎËüÝ(lĘói)ď┤´@Í°Ë░ÝĹĚN├š│÷═┴║═╔˙╬´┴┐Á╚╔˙ÚL(zhĘúng)▒ÝČF(xiĘĄn)�úČ▓╗Ş▓═┴Ď▓´@Í°ŻÁÁ═┴╦ĚN├š│÷═┴▓óĐËÚL(zhĘúng)┴╦│÷├šĽr(shʬ)Úgíúď┌▀M(jĘČn)đđÍ▓▒╗╗ÍĆ═(fĘ┤)╣▄└ÝîŹ(shʬ)█`Ľr(shʬ)���úČ▀xô˝║¤▀mÁ─ĚNÎËüÝ(lĘói)ď┤║═┐ěÍĂ▓ąĚN╔ţÂ╚╩ăĚă│ú▒ěϬÁ─���íú
7úęíó╠ý╚╗┐Ţ▓ŢîŽ(duĘČ)ĚN├šŻĘÍ▓┐éÁ─üÝ(lĘói)Ňf(shuĘş)Ëđěô(fĘ┤)├ŠË░ÝĹ����úČ͸Ϭ▒ÝČF(xiĘĄn)×ÚŻÁÁ═┴╦ĚNÎË│÷├š┬╩úČ╝┤═Ę▀^(guĘ░)ÎŔÍ╣ĚNÎË┼c═┴╚└ŻËË|°´@Í°ŻÁÁ═│÷├š┬╩�íúďç˛×(yĘĄn)▒Ý├¸╬╗Ë┌═┴╚└ÍđÁ─ĚNÎËĂń│÷├š┬╩´@Í°Ş▀Ë┌┐Ţ▓ŢîË▒Ý├Šíó┐Ţ▓ŢîË┼c═┴╚└▒Ý├ŠÁ─│÷├š┬╩���íúď┌╠ý╚╗▓ŢďşÍ▓▒╗╗ÍĆ═(fĘ┤)╣▄└ÝîŹ(shʬ)█`Íđ�úČ▓╔╚í▀m«ö(dĘíng)?shĘ┤)─┤Ű╩ę╚ą│ř┐Ţ▓Ţ╩╣ĚNÎË┬ń╚Ű═┴╚└����úČ▓┼─▄▒úÎCĚNÎËËđŢ^Ş▀Á─│÷├š┬╩║═ĚN├š│╔╣ŽŻĘÍ▓íú
8úęíóć╬▓ą║═╗ý▓ąÁ─îŹ(shʬ)˛×(yĘĄn)Â╝▒Ý├¸�����úČčo(wĘ▓)├ó╚ŞűťÁ─ĚN├šŻĘÍ▓─▄┴Ž║═Şé(jĘČng)áÄ(zhĘąng)┴ŽĎ¬â×(yĘşu)Ë┌в▓Ţ��íú
│§▓ŻŻY(jiĘŽ)Ňô╚š¤┬ú║
Ë░ÝĹв▓Ţ│÷├šÁ─͸Ϭϲ╦ě╩ăĚNÎË┼c═┴╚└Á─ŻËË|│╠Â╚║═▓ąĚN╔ţÂ╚�����úČŞ▓╔w┐Ţ▓ŢîËËđ└űË┌Ď░═Ôв▓ŢĚN├š┤Š╗ţ�����íú
îŽ(duĘČ)═┴╚└▀M(jĘČn)đđö_äË(dĘ░ng)���úČ░ĎÁěŠé(zhĘĘn)ë║Ďď╩╣ĚNÎË┼c═┴╚└│ńĚÍŻËË|úĘĚNÎË╚Ű═┴1-2cmúę╩ă╠ߪ▀ĚN├šË╚Ăń╩ăв▓ŢĚN├š│÷├š┬╩Á─▒ěϬŚl╝■úČĎ▓╩ăĐa(bĘ│)▓ąĐ˛▓Ţ│╔╣Ž╗ÍĆ═(fĘ┤)▓ŢÁěÍ▓▒╗Á─ŕP(guĘín)ŠI���ú╗ŻÁÁ═üÝ(lĘói)ÎďČF(xiĘĄn)┤ŠÍ▓▒╗Á─Şé(jĘČng)áÄ(zhĘąng)�����úČË╚Ăń╩ăÁě¤┬Á─Şé(jĘČng)áÄ(zhĘąng)╩ă▒úÎCв▓Ţ�����íóčo(wĘ▓)├ó╚ŞűťĚN├šŻĘÍ▓│╔╣ŽÁ─▒ěϬŚl╝■�íú
5íó└űË├ĚŻ╩ŻîŽ(duĘČ)╠ý╚╗▓ŢÁě─┴▓Ţ╔˙ÚL(zhĘúng)┼cĚ▒Í│Á─Ë░ÝĹĐđż┐
ď┌║Ë▒▒Ćł╝Ď┐┌ë╬╔¤╣┴ď┤─┴ł÷(chĘúng)в▓Ţ▓Ţďş╔¤�úČ▀xô˝Í▓▒╗ż¨ä˛íó╬óÁěđ╬▓ţ«ÉŢ^đíÁ─Şţ▓ŢÁě║═Ě┼─┴Áěθ×ÚĐđż┐śËÁě�����íúĎďĚN╚║×ÚĐđż┐îŽ(duĘČ)¤ˇ�úČË┌2005─ŕ6ď┬íź2005─ŕ10ď┬úČîŽ(duĘČ)─┴▓ŢÁ─Ě▒Í│îŽ(duĘČ)▓▀║═ĚNÎËËŕ▀M(jĘČn)đđ┴╦Đđż┐����úČĚÍ╬÷┴╦▓Ţďş─┴▓Ţď┌ĚN╚║╦«ĂŻ╔¤Á─Ě▒Í│ĎÄ(guĘę)┬╔║═Í▓▒╗ĐŢ╠ŠĎÄ(guĘę)┬╔íú
5.1 Ě┼─┴Áě┼cŞţ▓ŢÁěÍ▓▒╗Ň{(diĘĄo)▓Ú
Şţ▓ŢÁěâ×(yĘşu)äŢ(shĘČ)Â╚Îţ┤ˇÁ─ă░╬ňĚNÍ▓╬´ĚÍäe×Úú║в▓ŢúĘ59.6úę��íó▓┌Ű[ÎË▓ŢúĘ9.51úę���íó─¤─ÁŢ´úĘ3.80úę�íó└ńŢ´úĘ2.96úę�íóßś├ęúĘ2.74úęúČĂńÍđв▓ŢŇ╝ËđŻ^îŽ(duĘČ)â×(yĘşu)äŢ(shĘČ)�����ú╗°Ě┼─┴Áěâ×(yĘşu)äŢ(shĘČ)Â╚Îţ┤ˇÁ─ă░╬ňĚNÍ▓╬´ĚÍäe×Úú║в▓ŢúĘ34.4úęíó▓┌Ű[ÎË▓ŢúĘ9.36úę���íóßś├ęúĘ8.06úę����íóĂĐ╣źËóúĘ6.04úę����íó˝R╠AúĘ3.96úę�íúË╔Ë┌в▓ŢÁ─â×(yĘşu)äŢ(shĘČ)Â╚ŻÁÁ═úČĚ┼─┴ÁěÍđÁ─▓┌Ű[ÎË▓Ţ��íó─¤─ÁŢ´�����íóßś├ęíó╝Ü(xĘČ)┴Đ╚~Ţ´íó├╔╣┼Ţ´�íóÂ■┴Đ╬«┴ŕ▓╦íó╝Ü(xĘČ)╚~ż┬���íó˙Q╩şíóâ╔ŚźŮĄÁ╚╬´ĚNÁ─╔wÂ╚íó├▄Â╚║═â×(yĘşu)äŢ(shĘČ)Â╚Â╝┤ˇË┌Şţ▓ŢÁě═ČĚNÍ▓╬´Á─╔wÂ╚��íó├▄Â╚║═â×(yĘşu)äŢ(shĘČ)Â╚�����íú
▒Ý1-25 Ě┼─┴Áě���íóŞţ▓ŢÁěÍ▓╬´ĚNÁ─Ş▀Â╚��íó╔wÂ╚�íó├▄Â╚╝░â×(yĘşu)äŢ(shĘČ)Â╚Á─▒╚Ţ^
|
╬´ĚN
|
Şţ▓ŢÁě |
Ě┼─┴Áě |
|
Ş▀Â╚ ╔wÂ╚ ├▄Â╚ â×(yĘşu)äŢ(shĘČ)Â╚ |
Ş▀Â╚ ╔wÂ╚ ├▄Â╚ â×(yĘşu)äŢ(shĘČ)Â╚ |
|
в▓Ţ |
Leymus chinesis Tzvel. |
22.2 |
81 |
806 |
59.6 |
6.46 |
43 |
1244 |
34.4 |
|
▓┌Ű[ÎË▓Ţ |
Poa annua L. |
7.69 |
26.3 |
47 |
9.51 |
4.15 |
28 |
59 |
9.36 |
|
ßś├ę |
Stipa grandis P. |
11.1 |
4 |
5.9 |
2.74 |
7.45 |
17 |
66.7 |
8.06 |
|
╠Ž▓Ţ |
Carex cranaocarpa Mey. |
|
|
|
|
4.56 |
4.33 |
9.4 |
3.16 |
|
─¤─ÁŢ´ |
Artemisia eriopoda Bge. |
3.98 |
14.2 |
15 |
3.8 |
2.01 |
8 |
103.4 |
3.42 |
|
└ńŢ´ |
Artemisia feddei Levl. |
3.99 |
12.2 |
7.6 |
2.96 |
|
|
|
|
|
╝Ü(xĘČ)┴Đ╚~Ţ´ |
Artemisia tanacetifolia L. |
25.6 |
1.3 |
0.1 |
0.77 |
3.42 |
6 |
40.4 |
3.78 |
|
ĎŕÉŢ´ |
Artemisia capillaris Thunb. |
8.1 |
0.7 |
0.6 |
0.57 |
5.25 |
0.33 |
0.2 |
0.23 |
|
├╔╣┼Ţ´ |
Artemisia mongolica Fisch. |
4.4 |
1.0 |
0.9 |
0.45 |
2.72 |
2 |
1.9 |
1.26 |
|
ěi├źŢ´ |
Artemisia scoparia Wald. |
|
|
|
|
3.48 |
5 |
25.6 |
2.02 |
|
ĂĐ╣źËó |
Taraxacum mongolicum |
|
|
|
|
3.18 |
14.7 |
50.4 |
6.05 |
|
░óáľ╠ę╣Ě═Ů╗Ę |
Heteropappus altaicus Nov. |
9.4 |
1 |
0.5 |
0.91 |
4.90 |
0.33 |
0.2 |
0.30 |
|
┐ÓŢĄ▓╦ |
Ixeris sonchifolia Hance. |
|
|
|
|
3.80 |
0.67 |
0.3 |
0.26 |
|
╗Żq▓Ţ |
Leontopodium leontopodioides |
5.7 |
2.3 |
0.3 |
0.54 |
|
|
|
|
|
▓ŢÁě´L(fĘąng)├źżŇ |
Saussurea amara DC. |
|
|
|
|
2.64 |
5 |
14.6 |
1.71 |
|
Á╣Ë╚~´L(fĘąng)├źżŇ |
Saussurea runicinata DC. |
|
|
|
|
3.34 |
5 |
9.1 |
2.39 |
|
¨ZŻq╬«┴ŕ▓╦ |
Potentilla anserina L. |
|
|
|
|
3.23 |
3.33 |
34.4 |
2.71 |
|
Â■┴Đ╬«┴ŕ▓╦ |
Potentilla bifurca L. |
13.0 |
1.2 |
0.1 |
0.54 |
2.01 |
4.67 |
7.8 |
1.77 |
|
ÂÓăo╬«┴ŕ▓╦ |
Potentilla multicaulis Bge. |
17.4 |
2.7 |
0.7 |
2.37 |
3.8 |
1.67 |
0.1 |
0.45 |
|
ÁěË▄ |
Sanguisorba officinalis L. |
|
|
|
|
3.40 |
0.33 |
0.1 |
0.17 |
|
╔Żż┬(╔Ż╩[) |
Allium senescene L. |
9.6 |
1.7 |
0.7 |
1.03 |
6.78 |
0.67 |
1 |
0.55 |
|
╝Ü(xĘČ)╚~ż┬ |
Allium tenuissimum L. |
12.5 |
3.7 |
2.9 |
2.25 |
7.57 |
4.67 |
14.6 |
2.70 |
|
▒ÔŮúÂ╣ |
Medicago ruthenica L. |
4.8 |
3.3 |
0.7 |
1.0 |
3.00 |
3.33 |
1 |
1.66 |
|
Í▒┴óŘS▄╬ |
Astragalus adsurgens pall. |
25.6 |
1.2 |
0.1 |
0.91 |
3.17 |
3.67 |
0.9 |
2.12 |
|
┼űßś╚~ŘS╚A |
Thermopsis lanceolata R. Br. |
20.5 |
0.7 |
0.1 |
0.64 |
|
|
|
|
|
╚A▒▒ÄrŘS▄╬ |
Hedysarum gmelinii Ledeb. |
25.6 |
1.3 |
0.1 |
1.3 |
|
|
|
|
|
ŘS├ź╝ČÂ╣ |
Oxytropis ochrantha Bunge |
13.3 |
1.3 |
0.4 |
0.64 |
|
|
|
|
|
¬Ü(dĘ▓)đđ▓╦ |
Lepidium apetalum Willd. |
|
|
|
|
3.59 |
3.67 |
4.6 |
1.87 |
|
ăě▄┤ |
Gentiana macrophylla Pall. |
|
|
|
|
4.18 |
1.33 |
0.9 |
1.03 |
|
└ăÂż |
Stellera chamaejasme L. |
|
|
|
|
6.45 |
0.67 |
0.2 |
0.65 |
|
˝R╠A |
Iris ensata Thunb. |
|
|
|
|
11.0 |
5 |
1.9 |
3.96 |
|
┬űăo¤ëÎË▓Ţ |
Silene repens Patr. |
13.8 |
1.3 |
0.5 |
1.1 |
8.10 |
0.33 |
0.1 |
0.32 |
|
╝Ü(xĘČ)╚~Ă┼Ă┼╝{ |
Veronica linariifolia Pall. |
|
|
|
|
1.59 |
1 |
0.5 |
0.26 |
|
▀_(dĘó)ק└´đż░┼ |
Cymbaria dahurica L. |
4.7 |
1.3 |
1.5 |
0.47 |
|
|
|
|
|
▓˝║˙ |
Bupleurum chinensis DC. |
8.0 |
0.3 |
0.1 |
0.31 |
3.62 |
0.67 |
0.4 |
0.39 |
|
Ě└´L(fĘąng) |
Saposhmikovia divaricata S. |
7.47 |
9.2 |
0.3 |
1.2 |
|
|
|
|
|
ëA┼ţ |
Suaeda glauca Bge. |
|
|
|
|
5.85 |
2 |
1.7 |
1.06 |
|
Ď░║˙┬Ú |
Linum stelleroides L. |
17.0 |
0.7 |
0.3 |
0.59 |
|
|
|
|
|
â╔ŚźŮĄ |
Polygonum amphibium L. |
16.5 |
0.3 |
0.1 |
0.49 |
8.5 |
1.33 |
0.4 |
0.87 |
|
˙Q╩ş |
Lappula myosotis V. |
21.5 |
0.7 |
0.2 |
0.58 |
13.9 |
0.67 |
0.6 |
1.00 |
|
╠Ă╦╔▓Ţ |
Thalictrum petaloideum L. |
14.5 |
0.7 |
0.2 |
1.0 |
|
|
|
|
|
ëKăo▓┌╠K |
Phlomis umbrosa Turcz. |
13.0 |
2.3 |
0.4 |
1.5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
5.2 Ě┼─┴Áě┼cŞţ▓ŢÁěĚNÎËËŕ▒╚Ţ^
Şţ▓ŢÁěĚNÎËËŕ┴┐×Ú13968.6┴ú/ęO�����úČĚ┼─┴Áě×Ú1222.6┴ú/ęO�íúŞţ▓ŢÁěĚNÎËËŕÍđ║╠▒ż┐Ă║═żŇ┐ĂÍ▓╬´ĚNÎËŇ╝Ż^┤ˇÂÓöÁ(shĘ┤)úČĚÍäe×Ú31.7úą║═33.3úą����ú╗Ě┼─┴ÁěĚNÎËËŕ║╠▒ż┐Ă║═żŇ┐ĂÍ▓╬´ĚNÎËĎ▓Ň╝Ż^┤ˇ▓┐ĚÍúČĚÍäe×Ú36.3úą║═50.6úą���íúŞţ▓ŢÁěĚNÎËËŕ├▄Â╚Îţ┤ˇÁ─ă░╬ňĚNÍ▓╬´ĚÍäe×Úú║─¤─ÁŢ´���íó▓┌Ű[ÎË▓Ţ���íóĚ└´L(fĘąng)íó▓˝║˙║═в▓Ţ�íúĚ┼─┴ÁěĚNÎËËŕ├▄Â╚Îţ┤ˇÁ─ă░╬ňĚNÍ▓╬´ĚÍäe×Úú║▓┌Ű[ÎË▓ŢíóĂĐ╣źËó��íó─¤─ÁŢ´��íóв▓Ţ║═└ăÂż�úČŻY(jiĘŽ)ĚN┴┐Â╝đíË┌Şţ▓ŢÁěíú
▒Ý1-26 Ě┼─┴Áě���íóŞţ▓ŢÁěĚNÎËËŕ├▄Â╚
|
╬´ĚN |
╔˙╗ţđ═ |
ĚNÎËËŕ├▄Â╚úĘ┴ú/ęOúę |
|
Şţ▓ŢÁě |
Ě┼─┴Áě |
|
в▓Ţ |
Leymus chinesis Tzvel. |
PH |
1162.15 |
87.01 |
|
▓┌Ű[ÎË▓Ţ |
Poa annua L. |
PH |
3243.71 |
336.61 |
|
ßś├ę |
Stipa grandis P. |
PH |
20 |
17.5 |
|
▒Ô╦Ű▒¨▓Ţ |
Agropyron cristatum (L.) Gaertn |
PH |
6.2 |
2.1 |
|
╠Ž▓Ţ |
Carex cranaocarpa Mey. |
PH |
|
30.7 |
|
─¤─ÁŢ´ |
Artemisia eriopoda Bge. |
PH |
3690.7 |
265.25 |
|
└ńŢ´ |
Artemisia feddei Levl. |
SS |
5.4 |
0.28 |
|
╝Ü(xĘČ)┴Đ╚~Ţ´ |
Artemisia tanacetifolia L. |
PH |
28.2 |
11.76 |
|
ĎŕÉŢ´ |
Artemisia capillaries Thunb. |
PH |
500.2 |
|
|
├╔╣┼Ţ´ |
Artemisia mongolica Fisch. |
PH |
|
33.55 |
|
ěi├źŢ´ |
Artemisia scoparia Wald. |
ABH |
|
19.63 |
|
ĂĐ╣źËó |
Taraxacum mongolicum |
PH |
|
268.5 |
|
░óáľ╠ę╣Ě═Ů╗Ę |
Heteropappus altaicus Nov. |
PH |
29.4 |
11.6 |
|
┐ÓŢĄ▓╦ |
Ixeris sonchifolia Hance. |
PH |
|
7.5 |
|
╗Żq▓Ţ |
Leontopodium leontopodioides |
PH |
396.9 |
|
|
Â■┴Đ╬«┴ŕ▓╦ |
Potentilla bifurca L. |
PH |
53.8 |
|
|
ÂÓăo╬«┴ŕ▓╦ |
Potentilla multicaulis Bge. |
PH |
45.6 |
|
|
╔Żż┬(╔Ż╩[) |
Allium senescene L. |
PH |
261 |
|
|
╝Ü(xĘČ)╚~ż┬ |
Allium tenuissimum L. |
PH |
122.5 |
|
|
▒ÔŮúÂ╣ |
Medicago ruthenica L. |
PH |
111.3 |
4.7 |
|
Í▒┴óŘS▄╬ |
Astragalus adsurgens pall. |
PH |
21.9 |
|
|
┼űßś╚~ŘS╚A |
Thermopsis lanceolata R. Br. |
PH |
4.5 |
|
|
╚A▒▒ÄrŘS▄╬ |
Hedysarum gmelinii Ledeb. |
PH |
85.1 |
|
|
ŘS├ź╝ČÂ╣ |
Oxytropis ochrantha Turcz. |
PH |
3.1 |
|
|
¬Ü(dĘ▓)đđ▓╦ |
Lepidium apetalum Willd. |
AH |
|
14.4 |
|
đí╗Ę╗ĘĂýŚU |
Dontostemon micranthus C. A. Mey. |
AH |
108.8 |
|
|
ăě▄┤ |
Gentiana macrophylla Pall. |
PH |
|
|
|
└ăÂż |
Stellera chamaejasme L. |
PH |
30.3 |
74.3 |
|
˝R╠A |
Iris ensata Thunb. |
PH |
0.9 |
18.5 |
|
┬űăo¤ëÎË▓Ţ |
Silene repens Patr. |
ABH |
4.1 |
|
|
▀_(dĘó)ק└´đż░┼ |
Cymbaria dahurica L. |
PH |
19.4 |
|
|
▓˝║˙ |
Bupleurum chinensis DC. |
PH |
1814.4 |
|
|
Ě└´L(fĘąng) |
Saposhmikovia divaricata S. |
PH |
2113.4 |
|
|
ëA┼ţ |
Suaeda glauca Bge. |
AH |
|
1.5 |
|
Ď░║˙┬Ú |
Linum stelleroides L. |
ABH |
|
|
|
â╔ŚźŮĄ |
Polygonum amphibium L. |
PH |
4.9 |
|
|
˙Q╩ş |
Lappula myosotis V. |
AH |
35.7 |
17.2 |
|
░ŕ╚´╠Ă╦╔▓Ţ |
Thalictrum petaloideum L. |
PH |
7.9 |
|
|
ëKăo▓┌╠K |
Phlomis umbrosa Turcz. |
PH |
37.2 |
|
|
┐éËő(jĘČ)ú║ |
13968.66 |
1222.59 |
|
|
|
|
|
|
Îó: AH: Ď╗─ŕ╔˙▓Ţ▒ż; ABH: Ď╗Â■─ŕ╔˙▓Ţ▒ż: BH: Â■─ŕ╔˙▓Ţ▒ż; PH: ÂÓ─ŕ╔˙▓Ţ▒ż; SS: ░Ű╣Ó─ż(đí░Ű╣Ó─ż)
5.3 Ě┼─┴Áě┼cŞţ▓ŢÁěĚNÎËÄý(kĘ┤)▒╚Ţ^
Şţ▓ŢÁěíóĚ┼─┴Áě═┴╚└ĚNÎËÄý(kĘ┤)âŽ(chĘ│)┴┐Á─Ľr(shʬ)ÚgäË(dĘ░ng)ĹB(tĘĄi)ĚÍäe×Ú747.7íź1505.4┴ú/ęO║═303.0íź520.7┴ú/ęO��ú╗Şţ▓ŢÁěĚNÎËÄý(kĘ┤)ĚÍîËäË(dĘ░ng)ĹB(tĘĄi)úĘ┐ŇÚgäË(dĘ░ng)ĹB(tĘĄi)úęĚÍäe×Úú║0íź2ęM═┴îËĚNÎË├▄Â╚445.0íź833.2┴ú/ęO�����íó2íź5ęM═┴îËĚNÎË├▄Â╚170.4íź463.9┴ú/ęO║═5íź10ęM═┴îËĚNÎË├▄Â╚132.6íź208.3┴ú/ęO����úČĚ┼─┴ÁěĚNÎËÄý(kĘ┤)ĚÍîËäË(dĘ░ng)ĹB(tĘĄi)ĚÍäe×Úú║0íź2ęM═┴îËĚNÎË├▄Â╚179.9íź340.8┴ú/ęO��íó2íź5ęM═┴îËĚNÎË├▄Â╚85.2íź161.0┴ú/ęO║═5íź10ęM═┴îËĚNÎË├▄Â╚18.9íź28.4┴ú/ęO���íúŞţ▓ŢÁěíóĚ┼─┴ÁěĚNÎËÄý(kĘ┤)͸Ϭ╝»Íđď┌0íź5ęM═┴îËÍđ��úČË╔ť\╚Ű╔ţ°▀fťp�íúĚ┼─┴┐╔╩╣żŇ┐ĂÍ▓╬´ĚNÎË▒╚└řď÷┤ˇíúĚNÎËÄý(kĘ┤)ÍđżŇ┐ĂÍ▓╬´ĚNÎË░l(fĘí)Đ┐╦┘Â╚Ϭ┐ýË┌║╠▒ż┐Ă║═Â╣┐Ă��úČ╦¨ĎďďşËđÍ▓▒╗ĂĂë─Ďď║ˇ���úČżŇ┐ĂÍ▓╬´äŢ(shĘČ)▒ěϬŇ╝ţI(lĘźng)┐ŇÚg�����úČîž(dĘúo)Í┬─┴▓Ţ┘|(zhĘČ)┴┐¤┬ŻÁ��íú
┴Ý═ÔŞţ▓ŢîŽ(duĘČ)▓ŢÁěÍ▓╬´Á─Ş╔ö_│╠Â╚đíË┌Ě┼─┴îŽ(duĘČ)▓ŢÁěÍ▓╬´Á─Ş╔ö_���ú╗Ě┼─┴îž(dĘúo)Í┬Í▓Íŕđíđ═╗»íóިăo╣Ł(jiĘŽ)ÚgżÓŰx┐sÂ╠��ú╗═ČĽr(shʬ)úĘŢpÂ╚úęĚ┼─┴Ď▓┐╔Ďď╠ߪ▀▓ŢÁěÍ▓╬´ÂÓśËđď�����íú
6íó▓ŢÁě▓ŢÁěÍ▓▒╗îŽ(duĘČ)ŻÁËŕ┴┐Îâ╗»Á─ÝĹŬ(yĘęng)
▒żďç˛×(yĘĄn)Ë┌2005─ŕ6ď┬ď┌║Ë▒▒ë╬╔¤Ćł╝Ď┐┌╣┴ď┤─┴ł÷(chĘúng)в▓Ţ▓Ţďş�����úČ▀M(jĘČn)đđ┴╦ŻÁËŕ┴┐Á─▓ĘäË(dĘ░ng)║═└űË├ĆŐ(qiĘóng)Â╚Îâ╗»Á─ďç˛×(yĘĄn)╠Ä└Ý�����íúŻÁËŕ┴┐▓ĘäË(dĘ░ng)Á─╠Ä└Ý╩ă▓╔Ë├Ň┌ËŕďO(shĘĘ)éńörŻě50úąďç˛×(yĘĄn)đíů^(qĘ▒)0.5mŇř╔¤┐ŇÁ─├ŠĚe����úČĎď▀_(dĘó)ÁŻťp╔┘50úąŻÁ╦«┴┐Á─Ϭăˇ����úČďç˛×(yĘĄn)îŹ(shʬ)ŰHťy(cĘĘ)ÂĘÁ─örŻěđž╣űď┌43úąúş48úąÍ«ÚgúČď÷╝Ë50úą╠Ä└ÝÁ─ďç˛×(yĘĄn)đíů^(qĘ▒)▓╔Ë├╚╦╣ĄŁ▓╦«Á─ŮkĚĘ▀_(dĘó)ÁŻĎ¬ăˇ�����íúިô■(jĘ┤)ÜÔ¤ˇŇżÁ─ťy(cĘĘ)ÂĘŻY(jiĘŽ)╣ű���úČ▀M(jĘČn)đđŁ▓╦«���íú└űË├ĆŐ(qiĘóng)Â╚ďO(shĘĘ)Í├â╔éÇ(gĘĘ)╠ŢÂ╚���úČĎ╗éÇ(gĘĘ)╚ź─ŕěÎŞţ2┤╬úČĎ╗éÇ(gĘĘ)âHěÎŞţĎ╗┤╬úę�����íúîŽ(duĘČ)ďç˛×(yĘĄn)╠Ä└ÝÍđÁ─в▓ŢÁě╔¤Áě¤┬╔˙╬´┴┐����íóďç˛×(yĘĄn)╠Ä└Ý▓╗═ČîË┤╬═┴╚└║Č╦«┴┐─ŕÂ╚Îâ╗»íóв▓Ţ╚║┬ńŻM│╔Ďď╝░▓╗═ČŻMĚÍ«a(chĘún)┴┐Á─ĚÍ▓╝Á╚▀M(jĘČn)đđ┴╦Đđż┐��íú
6.1 ŻÁ╦«┴┐Îâ╗»║═└űË├ĆŐ(qiĘóng)Â╚îŽ(duĘČ)Áě╔¤╔˙╬´┴┐Îâ╗»Á─Ë░ÝĹ
ŰS└űË├ĆŐ(qiĘóng)Â╚Á─ď÷╝Ë���úČ▓ŢÁěÁ─Áě╔¤╔˙╬´┴┐ËđŻÁÁ═Á─┌ůäŢ(shĘČ)�úČď÷╝ËŻÁ╦«┴┐║═ťp╔┘ŻÁËŕ┴┐Â╝Ď╗ÂĘ│╠Â╚Á─ťp╔┘┴╦в▓Ţ╚║┬ńÁ─╔˙╬´┴┐Á─┌ůäŢ(shĘČ)�����úČĚŻ▓ţĚÍ╬÷▓╗´@Í°����íúěÎŞţĆŐ(qiĘóng)Â╚ď÷╝Ë═ČĽr(shʬ)ŻÁËŕ┴┐ťp╔┘Á─ăÚŤr¤┬�úČ╔˙╬´┴┐´@Í°ŻÁÁ═��ú╗ď┌Ňř│ú║═ď÷╝Ë50úąŻÁ╦«┴┐╠Ä└ÝÍđď÷╝Ë└űË├ĆŐ(qiĘóng)Â╚�����úČ╔˙╬´┴┐ŻÁÁ═▓ó▓╗├¸´@����íúŇf(shuĘş)├¸└űË├ĆŐ(qiĘóng)Â╚Á─ď÷╝ËĽ■(huĘČ)ťp╔┘Áě╔¤╔˙╬´┴┐úČË╚Ăń╩ăŞ╔║ÁÁ──ŕĚŢĽ■(huĘČ)▀M(jĘČn)Ď╗▓Ż╝ËäíÁě╔¤╔˙╬´┴┐Á─ťp╔┘úĘłD1-10úę���íú
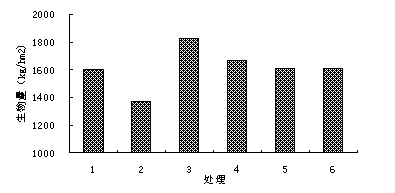
łD1-10 ▓╗═Č╠Ä└ÝÚg─ŕ╔˙╬´┴┐ÝĹŬ(yĘęng)úĘ2005─ŕúę
Îóú║╠Ä└Ý1▒Ý╩żŻÁËŕ┴┐ťp╔┘50úą��úČ╚ź─ŕěÎŞţ1┤╬����ú╗╠Ä└Ý2▒Ý╩żŻÁËŕ┴┐ťp╔┘50úą����úČ╚ź─ŕěÎŞţ2┤╬�ú╗╠Ä└Ý3▒Ý╩żŻÁËŕ┴┐▓╗ÎâúČ╚ź─ŕěÎŞţ1┤╬�����ú╗╠Ä└Ý4▒Ý╩żŻÁËŕ┴┐▓╗ÎâúČ╚ź─ŕěÎŞţ2┤╬��ú╗╠Ä└Ý5▒Ý╩żŻÁËŕ┴┐ď÷╝Ë50úą�úČ╚ź─ŕěÎŞţ1┤╬ú╗╠Ä└Ý6▒Ý╩żŻÁËŕ┴┐ď÷╝Ë50úą�úČ╚ź─ŕěÎŞţ2┤╬íú
6.2 ─ŕŰHÚgŻÁËŕ┴┐Îâ╗»îŽ(duĘČ)в▓Ţ╚║┬ńÁě╔¤╔˙╬´┴┐Ďď╝░╚║┬ńŻM│╔Á─Ë░ÝĹ
Ë╔łD1-11┐╔Ďď┐┤│÷2006─ŕÁ─╔˙ÚL(zhĘúng)╝żÁ─ŻÁ╦«┴┐×Ú311.79mm▒╚2005─ŕÁ─216.7ď÷╝Ë┴╦95.09mm.в▓Ţ╚║┬ńÁ─╔˙╬´┴┐├¸´@ď÷╝ËË╔2005─ŕÁ─159g/m2 ď÷╝ËÁŻ 191g/m2▀_(dĘó)ÁŻ´@Í°đď▓ţ«É���úČв▓ŢÁ─«a(chĘún)┴┐Ë╔112g/m2ťp╔┘ÁŻ100g/m2▀_(dĘó)ÁŻ´@Í°đď▓ţ«É�����íú
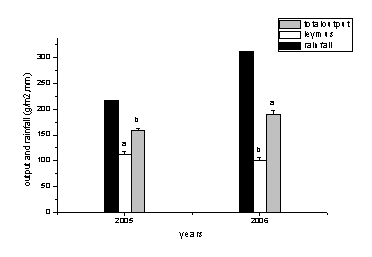
łD1-11 ŻÁËŕ┴┐║═Áě╔¤╔˙╬´┴┐─ŕÂ╚ÚgÎâ╗»
Ë╔2005║═2006─ŕ▓╗═Č╠Ä└ÝÚgв▓Ţ╚║┬ńŻĘ╚║ĚNŻM│╔Á─Îâ╗»┐╔Ďď┐┤│÷�����úČŰSÍ°ŻÁËŕ┴┐Á─ď÷╝Ë���úČď┌«a(chĘún)┴┐śő(gĘ░u)│╔╔¤─¤─ÁŢ´║═▓┌Ű[ÎË▓ŢÁ─▒╚└řď÷╝ËúČ°в▓ŢÁ─«a(chĘún)┴┐│╩ČF(xiĘĄn)¤┬ŻÁÁ─┌ůäŢ(shĘČ)����íúĚÍ╬÷͸ϬÁ─ďşĎ˛╩ă─¤─ÁŢ´║═▓┌Ű[ÎË▓ŢÁ─ި¤Á͸ϬĚÍ▓╝ď┌═┴╚└▒ÝîË0úş8cm▀@éÇ(gĘĘ)îË┤╬���úČв▓ŢÁ─ިăo╝»Íđď┌8úş14cmíúŻÁ╦«┴┐Á─ď÷╝Ë╠ߪ▀┴╦═┴╚└▒ÝîËÁ─║Č╦«┴┐��úČď÷╝Ë┴╦─¤─ÁŢ´║═▓┌Ű[ÎË▓Ţď┌ĚN╚║ÍđÁ─Şé(jĘČng)áÄ(zhĘąng)─▄┴Ž�����úČϲ┤╦в▓ŢÁ─╔˙╬´┴┐ď┌ŻÁ╦«┴┐ď÷╝ËÁ─ăÚŤr¤┬Ě┤°│÷ČF(xiĘĄn)┴╦¤┬ŻÁÁ─ČF(xiĘĄn)¤ˇúĘłD1-12��úČłD1-13úę����íú
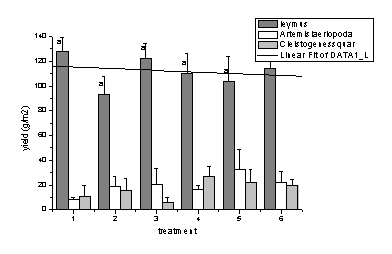
łD1-12 2005─ŕв▓Ţ╚║┬ńŻĘ╚║ĚNÁ─«a(chĘún)┴┐śő(gĘ░u)│╔
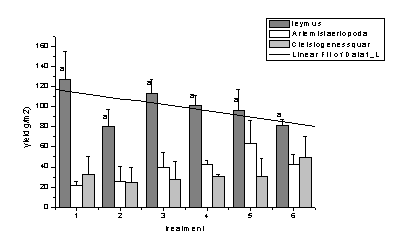
łD1-13 2006─ŕв▓Ţ╚║┬ńŻĘ╚║ĚNÁ─«a(chĘún)┴┐śő(gĘ░u)│╔
6.3└űË├ĆŐ(qiĘóng)Â╚îŽ(duĘČ)в▓ŢĚN╚║Á─Ë░ÝĹ
└űË├ĆŐ(qiĘóng)Â╚îŽ(duĘČ)в▓Ţ╚║┬ńŻĘ╚║ĚNÁ─Ë░ÝĹ║═ŻÁËŕ┴┐Á─┤ˇđíËđ├▄ăđÁ─ŕP(guĘín)¤ÁúČË╔łD1-14┐╔Ďď┐┤│÷���úČ2006─ŕŻÁ╦«┴┐ď÷╝Ë�úČ└űË├ĆŐ(qiĘóng)Â╚ď÷╝Ë║ˇĐ˛▓ŢÁ─╔˙╬´┴┐│╩ČF(xiĘĄn)ď÷╝ËÁ─┌ůäŢ(shĘČ)��ú╗°2005─ŕŻÁËŕ┴┐Ţ^Á═�úČŰS└űË├ĆŐ(qiĘóng)Â╚ď÷╝Ë║ˇúČв▓Ţ«a(chĘún)┴┐│÷ČF(xiĘĄn)¤┬ŻÁÁ─┌ůäŢ(shĘČ)�����úČď┌2005��íó2006─ŕŻÁËŕ┴┐Îâ╗»╩╣─¤─ÁŢ´║═▓┌Ű[ÎË▓ŢÁ─┐éÁě╔¤╔˙╬´┴┐Ë╔33úş40g/m2ď÷╝ËÁŻ72g/m2«a(chĘún)┴┐├¸´@ď÷╝Ë�ú╗°└űË├ĆŐ(qiĘóng)Â╚╠Ä└ÝîŽ(duĘČ)2005║═2006─ŕв▓Ţ╚║┬ńÍđÁ──¤─ÁŢ´║═▓┌Ű[ÎË▓ŢÁ─Ë░ÝĹ▓╗┤ˇúĘłD1-14úęíú
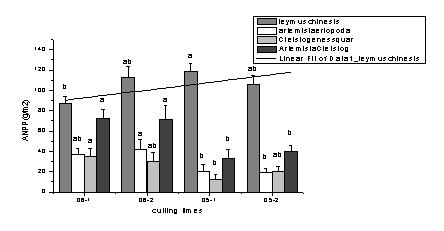
łD1-14 ěÎŞţĆŐ(qiĘóng)Â╚îŽ(duĘČ)в▓ŢĚN╚║ŻM│╔Á─║═Áě╔¤╔˙╬´┴┐Á─Ë░ÝĹ
6.4 ŻÁËŕ┴┐Îâ╗»îŽ(duĘČ)▓ŢÁě╔˙ÚL(zhĘúng)╦┘┬╩Á─Ë░ÝĹ
2006─ŕв▓Ţ╚║┬ńÁ─┐é«a(chĘún)┴┐▒╚2005─ŕ╠ߪ▀┴╦48.9úą�úČв▓Ţ«a(chĘún)┴┐╠ߪ▀43.9úąúČ2006─ŕ╔˙ÚL(zhĘúng)╝żŞ¸éÇ(gĘĘ)Ľr(shʬ)Ă┌Á─Áě╔¤╔˙╬´┴┐Â╝Ş▀Ë┌2005─ŕÁ─¤ÓŬ(yĘęng)Áě╔¤╔˙╬´┴┐���íú2006─ŕ╔˙ÚL(zhĘúng)╝żÁ─ŻÁËŕ┴┐×Ú318mm����úČď┌8ď┬20╚Ňă░ŻÁËŕ▒╚Ţ^ż¨ä˛���úČĆ─łDÍđ┐╔Ďď┐┤│÷2006─ŕв▓Ţ╚║┬ń┐é╔˙╬´┴┐║═в▓ŢÁ─╔˙╬´┴┐Â╝╩ăÍŁuď÷╝ËÁ─����úČ°2005─ŕ╔˙ÚL(zhĘúng)╝żÁ─ŻÁËŕ×Ú200mm°ăĎ╔˙ÚL(zhĘúng)╝żă░Ă┌Ăź║Á���úČ╔˙╬´┴┐Á─Ěe└█┤Šď┌├¸´@Á─═úÍ╣��úČĚ┤ĆŚÁ─ČF(xiĘĄn)¤ˇúĘłD1-15úę�íú
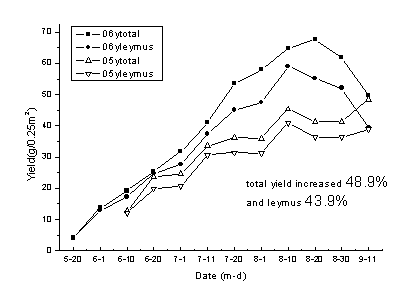
łD 1-15 2005─ŕ║═2006─ŕ▓╗═ČĽr(shʬ)Ă┌в▓Ţ┼c╚║┬ń┐é«a(chĘún)┴┐Á─Áě╔¤╔˙╬´┴┐Îâ╗»
îŽ(duĘČ)2006─ŕ╔˙ÚL(zhĘúng)╦┘┬╩úĘłD1-16úę▀M(jĘČn)đđĚÍ╬÷▒Ý├¸úČ8ď┬20╚Ňă░ŇűéÇ(gĘĘ)╚║┬ń╩ăŇřď÷ÚL(zhĘúng)�úČ8ď┬20╚ŇΡËĎ╚║┬ńÁ─Áě╔¤╔˙╬´┴┐▀_(dĘó)ÁŻĚňÍÁúČŻËÍ°╚║┬ńÁ─Áě╔¤╔˙╬´┴┐Ú_╩╝¤┬ŻÁ��íúÍÁÁ├ÎóĎÔÁ─╩ă8ď┬1╚Ň▀@Ď╗Řc(diĘún)��úČĆ─łDÍđ┐╔Ďď┐┤│÷▀@Ď╗Â╬Ľr(shʬ)Úg╔˙ÚL(zhĘúng)╦┘┬╩├¸´@ŻÁÁ═����úČĆ─ďç˛×(yĘĄn)ÁěÍđĐŰÁ─ďç˛×(yĘĄn)ŇżÁ├ÁŻÁ─ÜÔ¤ˇöÁ(shĘ┤)ô■(jĘ┤)╩ăŻÁËŕ┴┐34.4mmúČĂŻż¨ťěÂ╚17.7íŠ���úČ╠źŕľ(yĘóng)ŢŚ╔ń183.3w/m2�����úČ´L(fĘąng)╦┘6.4m/s�����úČ═┴╚└║Č╦«┴┐13.0úą����ú╗7ď┬20╚ŇÁ─öÁ(shĘ┤)ô■(jĘ┤)ĚÍäe×Ú66.3����íó19.4�íó242.3�íó73.9íó7.3��íó14.3��ú╗8ď┬10╚ŇÁ─×Ú27.5����íó20.0�����íó233.2���íó84.4�����íó5.0��íó11.0���í��ú┐╔Ďď┐┤│÷͸ϬÁ─ů^(qĘ▒)äeď┌┼cĂŻż¨ťěÂ╚����íó╠źŕľ(yĘóng)ŢŚ╔ń║═¤ÓîŽ(duĘČ)Ł˝Â╚�����úČË╔Ë┌▀@Â╬Ľr(shʬ)Úg╠źŕľ(yĘóng)ŢŚ╔ńÁ═�����úČҢ░l(fĘí)┴┐đí┐ŇÜÔÁ─¤ÓîŽ(duĘČ)Ł˝Â╚Ţ^Ş▀����úČ┐ŇÜÔťěÂ╚Ţ^Á═úČϲ┤╦╔˙ÚL(zhĘúng)╦┘┬╩ťpżĆ�íúË╔┤╦┐╔ĎŐ╝┤╩╣ď┌╔˙ÚL(zhĘúng)═˙╩óÁ─╝ż╣Ł(jiĘŽ)úČď┌░ŰŞ╔║ÁÁěů^(qĘ▒)ŻÁËŕ┴┐Ď▓▓╗╩ă╬ĘĎ╗ŤQÂĘϲ╦ě����úČ▀@Ď╗░l(fĘí)ČF(xiĘĄn)ÚgŻËÁ─╗ě┤┴╦×Ú╩▓├┤ď┌Ë├ŻÁËŕ┴┐┼c╔˙╬´┴┐▀M(jĘČn)đđ¤ÓŕP(guĘín)ĚÍ╬÷Ľr(shʬ)úČ▓ţ«É▓ó▓╗Ď╗ÂĘ´@Í°Á─ćľ(wĘĘn)ţ}����íú
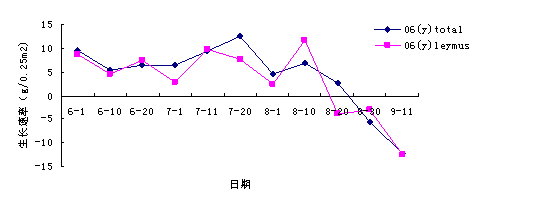
łD1-16 ▓╗═ČĽr(shʬ)Ă┌в▓Ţ║═╚║┬ńÁ─╔˙ÚL(zhĘúng)╦┘┬╩
6.5 ŻÁ╦«┴┐Îâ╗»║═└űË├ĆŐ(qiĘóng)Â╚îŽ(duĘČ)Áě¤┬╔˙╬´┴┐Á─Ë░ÝĹ
ďç˛×(yĘĄn)╠Ä└ÝÍđŰSÍ°ŻÁËŕ┴┐Á─ď÷╝ËÁě¤┬╔˙╬´┴┐Ëđťp╔┘Á─┌ůäŢ(shĘČ)úĘłD1-17úę���úČŰSÍ°ěÎ

łD1-17 ŻÁËŕ┴┐║═└űË├ĆŐ(qiĘóng)Â╚îŽ(duĘČ)Áě¤┬╔˙╬´┴┐Á─Ë░ÝĹ
Îóú║BNPP▒Ý╩żÁě¤┬╔˙╬´┴┐úČrhizome▒Ý╩żĐ˛▓ŢÁě¤┬ިăo���úČroots▒Ý╩żĂń╦ŘŞ¨¤Á����íú
Şţ┤╬öÁ(shĘ┤)Á─ď÷╝Ëď┌ťp╔┘50úąÁ─╠Ä└Ý║═ŻÁËŕ┴┐▓╗ÎâÁ─╠Ä└ÝÍđÁě¤┬╔˙╬´┴┐Ëđď÷╝ËÁ─┌ůäŢ(shĘČ)���íúď┌2005─ŕ╔┘Ëŕ─ŕ║═2006─ŕěSËŕ─ŕÁ─▒╚Ţ^ÍđúČ2006─ŕÁ─Í▓╬´Ş¨¤Á┐é┴┐ϬŞ▀Ë┌2005─ŕÁ─╦«ĂŻ���úČ▀@Ď╗ŻY(jiĘŽ)╣ű║═─ŕâ╚(nĘĘi)Á─ŻÁËŕ┴┐Îâ╗»Ňř║├¤ÓĚ┤��úČď┌ĚŻ▓ţĚÍ╬÷Íđ▓ţ«É╬┤▀_(dĘó)ÁŻ´@Í°╦«ĂŻ�íú
6.6 đíŻY(jiĘŽ)
ď┌░ŰŞ╔║Áů^(qĘ▒)▓Ţďş���úČθ×Ú͸ϬÎď╚╗¤ŮÍĂϲÎËÁ─ŻÁ╦«┴┐îŽ(duĘČ)в▓Ţ╚║┬ńÁ─Áě╔¤╔˙╬´┴┐Ë░ÝĹ║▄┤ˇ��úČ▒żďç˛×(yĘĄn)Íđ2006─ŕ┐é«a(chĘún)┴┐▒╚2005─ŕď÷╝Ë┴╦48.9úą��úČϲ┤╦╬Ďéâ▓╗─▄ĎďéÇ(gĘĘ)äe─ŕĚŢ▓Ţďş╔˙╬´┴┐ŻÁÁ═θ×Ú┼đÂĘ▓Ţł÷(chĘúng)═╦╗»Á─ÍŞś╦(biĘío)��íúϲ?yĘĄn)Úď┌Îď╚╗Śl╝■¤┬����úČ─ŕŰHÚg╩▄ŻÁËŕ┴┐Á─Ë░ÝĹúČ▓Ţł÷(chĘúng)╔˙╬´┴┐┤Šď┌║▄┤ˇÁ─▓ĘäË(dĘ░ng)đď�íú
ďç˛×(yĘĄn)ŻY(jiĘŽ)╣űĎ▓Ňf(shuĘş)├¸╬Ďéâ?cĘĘ)┌î?duĘČ)▓Ţďş▀M(jĘČn)đđ╣▄└ÝĽr(shʬ)úČ╚šď┌▀M(jĘČn)đđ▓ŢđˇĂŻ║Ô���úČĎď▓ŢÂĘđˇĽr(shʬ)đŔϬ┐╝Ĺ]Îď╚╗Śl╝■Á─Îâ╗»����úČË╚Ăń╩ăď┌░ŰŞ╔║Á▓Ţďşů^(qĘ▒)��íú
ď┌░ŰŞ╔║ÁÁěů^(qĘ▒)��úČÍ▓╬´═˙╩ó╔˙ÚL(zhĘúng)Á─Ľr(shʬ)Ă┌╚š╣ű╠źŕľ(yĘóng)ŢŚ╔ń║═ťěÂ╚ŻÁÁ═îóŻÁÁ═─┴▓ŢÁ─╔˙ÚL(zhĘúng)╦┘┬╩��íó¤ŮÍĂ▓ŢÁěÁ─╔˙╬´┴┐Á─Ěe└█�����íú
7ú«═╦╗»▓ŢÁě╗ÍĆ═(fĘ┤)Ş─┴╝─ú╩Ż
Ş╩├C▓Ţďş╔˙ĹB(tĘĄi)Đđż┐╦¨ĎďŮr(nĘ«ng)śI(yĘĘ)▓┐▓ŢÁěŮr(nĘ«ng)śI(yĘĘ)╔˙ĹB(tĘĄi)îW(xuĘŽ)ÍěŘc(diĘún)Ú_Ě┼îŹ(shʬ)˛×(yĘĄn)╩ĎúĘ23260271úę║═Ĺcŕľ(yĘóng)ŘS═┴Ş▀ďş▓ŢÁěŮr(nĘ«ng)śI(yĘĘ)¤ÁŻy(tĘ»ng)ďç˛×(yĘĄn)Ňż×ÚĎ└═đ����úČË┌2002---2006─ŕď┌Ş╩├Cşh(huĘón)┐h▀M(jĘČn)đđ┴╦═╦╗»▓ŢÁěżC║¤Ş─┴╝╝╝đg(shĘ┤)Đđż┐��úČßśîŽ(duĘČ)▓╗═Č═╦╗»ŰAÂ╬╠ý╚╗▓ŢÁě╠ß│÷┴╦▓╗═ČÁ─Ş─┴╝╝╝đg(shĘ┤)����íú
╠ß│÷┴╦▀mĎ╦Ë┌ÍěÂ╚═╦╗»▓ŢÁěÁ─í░ĚÔËř+äŁĂĂ+Đa(bĘ│)▓ąí▒żC║¤Ş─┴╝���íúîŽ(duĘČ)▒╚ďç˛×(yĘĄn)▒Ý├¸���úČĚÔËříóĐa(bĘ│)▓ą���íóäŁĂĂ▓ŢĂĄż¨─▄ë˛╩╣═╦╗»▓ŢÁěÁ├Ďď╗ÍĆ═(fĘ┤)úČÁź3ĚNĚŻĚĘżC║¤╝»│╔║ˇ���úČż▀Ëđ║ć(jiĘún)ć╬����íóĎÎ▓┘θ����íóđž╣ű├¸´@Á╚╠ěŘc(diĘún)�úČăđîŹ(shʬ)┐╔đđ����úČ▀mË┌ďç˛×(yĘĄn)ů^(qĘ▒)═ĂĆVíú
╠ß│÷┴╦▀mË┌ÍđÂ╚═╦╗»▓ŢÁěÁ─í░â╔╝żäŁů^(qĘ▒)Ţć─┴í▒Ş─┴╝╝╝đg(shĘ┤)�íú░Đ▓ŢÁěĚÍ×Ú┼»╝żĚ┼─┴ů^(qĘ▒)║═└ń╝żĚ┼─┴ů^(qĘ▒)â╔ţÉúČĚÍäeď┌└ń�����íó┼»â╔╝żäŁů^(qĘ▒)Ţć─┴����úČżC║¤═┴íó▓Ţ�íóđˇ╠ěҸúČ┤_ÂĘ▀mĎ╦Á─Ţdđˇ┴┐��úČ▓óË├í░Úô¤ŮŰpϲÎËĚĘí▒ÍŞîž(dĘúo)޸Ţć─┴đíů^(qĘ▒)Á─Ě┼─┴Ú_╩╝║═ŻY(jiĘŽ)╩°Ľr(shʬ)Úg��íúďô─ú╩Ż�úČ╝╚═Ę▀^(guĘ░)╝ĎđˇĚ┼─┴▒ú│ÍŇř│úÁ─▓ŢđˇŕP(guĘín)¤ÁúČżSÎo(hĘ┤)▓ŢÁěŻí┐ÁúČËÍ┐╔Ďď┼cÂ╠Ă┌Ě╩ËřŻY(jiĘŽ)║¤╔˙«a(chĘún)â╔╝żđˇ«a(chĘún)ĂĚúČŞ─╔ĂĎď═¨╝ĎđˇĚ╩Ëř͸Ϭ╝»ÍđË┌└ń╝żÁ─ć╬Ď╗─ú╩Ż����íú
╠ß│÷┴╦í░═╦ŞűÁěĚÔËří▒Á─Îď╚╗╗ÍĆ═(fĘ┤)╝╝đg(shĘ┤)��íú═╦ŞűÁ쯍(jĘęng)▀^(guĘ░)Şűθ�����úČ╦«ĚÍŚl╝■║├��úČ═┴╚└Ě╩┴ŽŞ▀���úČ«a(chĘún)▓Ţ┴┐ď┌╗ÍĆ═(fĘ┤)│§Ă┌║▄Ş▀�����úČ╬ŘĎř╝ĎđˇĚ┼─┴���úČÁź│§Ă┌͸Ϭ╔˙ÚL(zhĘúng)Ď╗─ŕ╔˙─┴▓Ţ�����úČĚ┼─┴îž(dĘúo)Í┬─┴▓Ţ╔˙ÚL(zhĘúng)▓╗│ńĚÍ��úČŰyĎďđ╬│╔Ě▒Í│ø╣┘úČ═┴ÁěĐŞ╦┘═╦╗»��úČ▓╔Ë├ĚÔËř║ˇ���úČ╩«ÄÎ─ŕż═┐╔ĎďĐŞ╦┘╗ÍĆ═(fĘ┤)ÁŻŻí┐Á╦«ĂŻ��íú
ďç˛×(yĘĄn)ů^(qĘ▒)╠ý╚╗▓ŢÁěÍ▓▒╗╔wÂ╚╗ÍĆ═(fĘ┤)ÁŻ90%Ďď╔¤��úČÁě╔¤╔˙╬´┴┐╠ߪ▀1▒ÂΡËĎ�íúď┌╔¤╩÷╝╝đg(shĘ┤)Á─Ížô╬¤┬���úČşh(huĘón)┐h╠ý╚╗▓Ţﺪ─┴╝╩żĚÂ┼c═ĂĆV├ŠĚe│Č▀^(guĘ░)2600hm2��íú═Ę▀^(guĘ░)Đa(bĘ│)▓ą���úČäŁĂĂÁ╚ĚŻ╩Ż╗ÍĆ═(fĘ┤)╠ý╚╗▓Ţďş500 hm2Ďď╔¤í�ú╗ÍĆ═(fĘ┤)ů^(qĘ▒)Í▓▒╗Ş▓╔w┬╩▀_(dĘó)ÁŻ75%Ďď╔¤úČ╩żĚÂů^(qĘ▒)═┴╚└ËđÖC(jĘę)┘|(zhĘČ)║Č┴┐ĂŻż¨╠ߪ▀0.2%Ďď╔¤�íúŢdđˇ┴┐3600вć╬╬╗úČÍ▄▀ůÓl(xiĘíng)Šé(zhĘĘn)300ÂÓŮr(nĘ«ng)ŢÍ▒ŻË╩▄ĎŠ����íú
7.1 í░ĚÔËř+äŁĂĂ+Đa(bĘ│)▓ąí▒żC║¤Ş─┴╝╝╝đg(shĘ┤)
ď┌ÍěÂ╚═╦╗»Á─ÎďË╔Ě┼─┴▓ŢÁě╔¤ŻĘ┴óç˙ĚÔ�����íóäŁĂĂ+ç˙ĚÔ�íóç˙ĚÔ+Đa(bĘ│)▓ą�����íóç˙ĚÔ+Đa(bĘ│)▓ą+äŁĂĂďç˛×(yĘĄn)śËÁě�����íúťy(cĘĘ)ÂĘďç˛×(yĘĄn)ă░║ˇŞ¸╠Ä└ÝÁ─Áě╔¤ČF(xiĘĄn)┤Š┴┐���íóÁ˛┬ń╬´║═╬´ĚNöÁ(shĘ┤)Á╚Í▓▒╗╠ěҸ�íú▒╚Ţ^޸ĚNŞ─┴╝┤Ű╩ęÁ─â×(yĘşu)Řc(diĘún)║═╚▒Řc(diĘún)�����úČ▓óîŽ(duĘČ)ŕP(guĘín)ŠI╝╝đg(shĘ┤)Ú_Ň╣îúÝŚ(xiĘĄng)Đđż┐�����íú
7.1.1 îŽ(duĘČ)▓ŢÁěÁě╔¤ČF(xiĘĄn)┤Š┴┐����íóÁ˛┬ń╬´┼c═┴╚└ËđÖC(jĘę)╠╝Á─Ë░ÝĹ
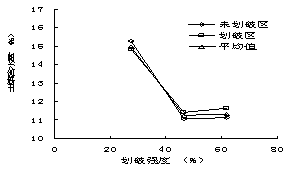
┐éˇw╔¤úČĚÔËřÚ_╩╝║ˇ1─ŕâ╚(nĘĘi)����úČÍěÂ╚═╦╗»▓ŢÁěÁě╔¤ČF(xiĘĄn)┤Š┴┐íóÁ˛┬ń╬´┼c┴ó┐Ţ╬´┴┐╝▒äíťp╔┘�úČ͸Ϭϲ?yĘĄn)ÚĎ╗─ŕ╔˙Í▓╬´ťp╔┘Ţ^ÂÓúČÁź═┴╚└ËđÖC(jĘę)╠╝ď┌5─ŕâ╚(nĘĘi)ÍŁuď÷╝ËúĘłD1-18úę�íú╚╗°úČ─│đęÁěđ╬Śl╝■¤┬��úČ╚šŕľ(yĘóng)Ă┬15íŃ║═45íŃ�úČĚÔËř─▄Ě˝´@Í°ď÷╝Ë▓ŢÁěÁě╔¤╔˙╬´┴┐úČ▀Ç╚íŤQË┌ŻÁ╦«Śl╝■í�����úČF(xiĘĄn)┤Š┴┐Á─ď÷̨����úČŕÄĂ┬ÎţŞ▀×Ú25%úČĂ┬Ýö×Ú20%í¬60%�úČŕľ(yĘóng)Ă┬▓╗│Č▀^(guĘ░)35%úČ╚řéÇ(gĘĘ)Ă┬¤˛Á─Îâ╗»┌ůäŢ(shĘČ)×ÚĂ┬Ýö>ŕľ(yĘóng)Ă┬>ŕÄĂ┬�íúŕÄĂ┬15íŃ║═ŕľ(yĘóng)Ă┬45íŃŚl╝■¤┬�����úČĚÔËřîŽ(duĘČ)«a(chĘún)▓Ţ┴┐Ť](mĘŽi)Ëđ´@Í°Ë░ÝĹúĘp>0.05úę���íúĚÔËř║ˇúČ35%-50%Á─Áě╔¤╬´┘|(zhĘČ)θ×ÚÁ˛┬ń╬´┤Šď┌Ë┌▓ŢÁěÍđ�íú▓ŢÁěÁ˛┬ń╬´┴┐îŽ(duĘČ)Ë┌▒úÎo(hĘ┤)═┴╚└úČťpżĆ╦«═┴┴¸╩žËđÍěϬθË├�����úČĚÔËř║ˇ▓ŢÁěÁ˛┬ń╬´ČF(xiĘĄn)┤Š┴┐▒╚╬┤ĚÔËř▓ŢÁě´@Í°ď÷╝ËúĘp<0.01úę���íú┼cĚ┼─┴Ă┬Áě¤Ó▒╚�úČď┌ŕÄĂ┬��úČ15íŃ�íó30íŃ║═45íŃÁ─ĚÔËřĂ┬ÁěÁ˛┬ń╬´┴┐ď÷╝Ë2-4▒ÂúČĂ┬Ýöät╠ߪ▀3-15▒Â��úČŕľ(yĘóng)Ă┬Ş▀4-15▒Â���íúď┌Ţ^×ÚŞ╔ď´Á─ŕľ(yĘóng)Ă┬║═Ă┬Ýö�úČĚÔËřîŽ(duĘČ)Ë┌Á˛┬ń╬´Ěe└█ËđŞŘ×ÚĚeśOÁ─θË├íú
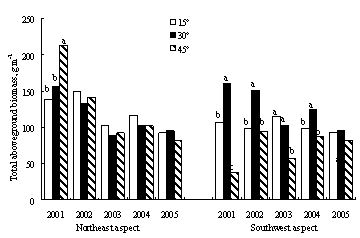

łD1-18ĚÔËř▓ŢÁěČF(xiĘĄn)┤Š┴┐���íóÁ˛┬ń╬´┴┐┼c═┴╚└ËđÖC(jĘę)╠╝║Č┴┐
7.1.2 îŽ(duĘČ)▓ŢÁě╔˙╬´┴┐íó╔wÂ╚║═╬´ĚNěSŞ╗Â╚Á─Ë░ÝĹ
޸ĚNŞ─┴╝┤Ű╩ę¤┬�����úČ─┴▓Ţ╔˙«a(chĘún)┴Ž┤ˇđíĎ└┤╬×Úú║äŁĂĂ+Đa(bĘ│)▓ą+ĚÔËř>Đa(bĘ│)▓ą+ĚÔËř>ĚÔËř>ÎďË╔Ě┼─┴úĘłD1-19úę�íúĚÔËř+Đa(bĘ│)▓ą+äŁĂĂ╠Ä└ÝÁ─▓ŢÁěúČ─┴▓ŢÁě╔¤ČF(xiĘĄn)┤Š┴┐úĘ225g/m2úę┼cÁ˛┬ń╬´┴┐úĘ54.93g/m2úęÎţŞ▀��úČĚÍäe╩ăĚÔËř▓ŢÁěÁ─1.9▒Â║═10▒Â���íúď┌Ě┼─┴Śl╝■¤┬��úČ┼cÍ▒ŻËĐa(bĘ│)▓ąÁ─Ş─┴╝ĚŻĚʤÓ▒╚����úČäŁĂĂ+Đa(bĘ│)▓ą╠Ä└ÝÁ─▓ŢÁě─┴▓ŢČF(xiĘĄn)┤Š┴┐┼cÁ˛┬ń╬´┴┐ĚÍäeď÷╝Ë0.43▒Â���íó6.66▒Â��íúŇf(shuĘş)├¸�úČ═╦╗»▓ŢÁěÁ─Ş─┴╝┼cĚ┼─┴▓ó▓╗îŽ(duĘČ)┴óíúÂ╠Ă┌â╚(nĘĘi)�úČ▓╗═Č╗ÍĆ═(fĘ┤)ĚŻ╩ŻîŽ(duĘČ)▓ŢÁě╬´ĚNöÁ(shĘ┤)íó╔wÂ╚Ë░ÝĹËđ╦¨▓ţ«É���úČŞ▀Ë┌ÎďË╔Ě┼─┴Áě20%-30%úĘłD1-19úę��íúŇf(shuĘş)├¸ĚÔËř─▄ë˛╠ߪ▀▓ŢÁěÁ─╔˙ĹB(tĘĄi)đžĎŠ�íúÁź╩ă�úČĚÔËř▓ó▓╗´@Í°ď÷╝Ë╬´ĚNěSŞ╗Â╚úĘp>0.05úęíú
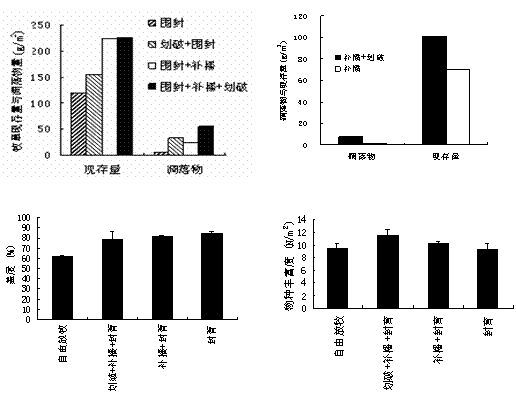
łD1-19 2004─સĚN╗ÍĆ═(fĘ┤)┤Ű╩ęîŽ(duĘČ)▓ŢÁě╔˙╬´┴┐��íó╔wÂ╚║═╬´ĚNěSŞ╗Â╚Á─Ë░ÝĹ
łD1-20 äŁĂĂĆŐ(qiĘóng)Â╚îŽ(duĘČ)▓ŢÁěÁ─Ë░ÝĹ
äŁĂĂĆŐ(qiĘóng)Â╚úĘäŁĂĂů^(qĘ▒)˲Ň╝ŇűéÇ(gĘĘ)▓ŢÁěÁ─├ŠĚe▒╚└řúę×Ú46.3%Ľr(shʬ)Áě╔¤╔˙╬´┴┐ÎţŞ▀úĘłD1-20úę����íúÁź╩㪨┴┐ď┌▒żÝŚ(xiĘĄng)─┐Á─äŁĂĂĆŐ(qiĘóng)Â╚ĚÂç˙â╚(nĘĘi)úČ┼cÍ«│╔Ňř▒╚��ú╗═┴╚└║Č╦«┴┐ŰSÍ«¤┬ŻÁ�����úČÁźäŁĂĂĆŐ(qiĘóng)Â╚46.3%┼c61.9%Í«Úgčo(wĘ▓)´@Í°▓ţ«ÉúĘp>0.05úę�����íúżC║¤3ÝŚ(xiĘĄng)ÍŞś╦(biĘío)úČĎďäŁĂĂĆŐ(qiĘóng)Â╚▓╗│Č▀^(guĘ░)50%×ÚĎ╦�úČ╚š┤╦úČ▓ŢÁě╝╚ż▀ËđŢ^Ş▀Á─Áě╔¤╔˙╬´┴┐��úČĎ▓ż▀ËđĎ╗ÂĘÁ─Áě¤┬╔˙╬´┴┐�����íú
▓╗═Čç˙ĚÔĽr(shʬ)¤ŮîŽ(duĘČ)▓ŢÁě╔˙«a(chĘún)┴Ž║═╬´ĚNěSŞ╗Â╚Ëđ´@Í°Ë░ÝĹúĘłD1-21úę�íú2005─ŕ�úČç˙ĚÔ6─ŕÁ─▓ŢÁě«a(chĘún)▓Ţ┴┐▒╚ÎďË╔Ě┼─┴ÁěŞ▀78.7%úĘp<0.01úęúČ2006─ŕç˙ĚÔ3─ŕ║═6─ŕÁ─śËÁě«a(chĘún)
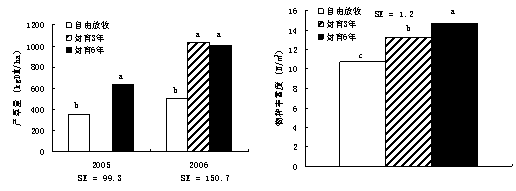
łD1-21 Ě┼─┴║═ĚÔËřîŽ(duĘČ)«a(chĘún)▓Ţ┴┐║═╬´ĚNěSŞ╗Â╚Á─Ë░ÝĹ
▓Ţ┴┐│Č│÷ÎďË╔Ě┼─┴Áě1▒ÂúĘp<0.01úę���íú2006─ŕ��úČ╬´ĚNěSŞ╗Â╚ĎďĚÔËř6─ŕÁ─śËÁěÎţŞ▀��úČ°ĎďÎďË╔Ě┼─┴ÁěÎţÁ═���íúŇf(shuĘş)├¸úČ▓ŢÁě╔˙«a(chĘún)┴ŽÁ─╗ÍĆ═(fĘ┤)Ϭ┐ýË┌╬´ĚNŻM│╔�íú
7.2 í░â╔╝żäŁů^(qĘ▒)Ţć─┴í▒Ş─┴╝╝╝đg(shĘ┤)
▀xô˝ÁěäŢ(shĘČ)ĂŻ╠╣íóÍ▓▒╗Ď╗Í┬Á─ÍđÂ╚═╦╗»╠ý╚╗▓ŢÁě�����úČŻĘ┴ó×ęвäŁů^(qĘ▒)Ţć─┴ďç˛×(yĘĄn)đíů^(qĘ▒)20éÇ(gĘĘ)íú¤─╝żŢć─┴▓ŢÁě�úČ├┐─ŕ6ď┬ÍđĐ«Í┴9ď┬ÍđĐ«Ţć─┴íú├┐đíů^(qĘ▒)▀B└m(xĘ┤)Ě┼─┴10d����úČŢć─┴Í▄Ă┌30dúČĎ╗─ŕŢć─┴3┤╬���íúďç˛×(yĘĄn)╝Ďđˇ×Ú1-2ÜqÁ─╚ąäŢ(shĘČ)╣ź×ęв����úȡwŤr¤ÓŻŘ���úČĚ┼─┴ĆŐ(qiĘóng)Â╚ĚÍäe×Ú0úĘîŽ(duĘČ)ŇŇúę�����íó2.67в/hm2���íó5.33в/ hm2║═8.67в/hm2íú├┐đíů^(qĘ▒)Ě┼─┴ă░úČ╝Ďđˇ┐ŇŞ╣ĚQÍě����úČ10╠ýĚQĎ╗┤╬íúĚ┼─┴ă░║ˇťy(cĘĘ)ÂĘŞ¸đíů^(qĘ▒)─┴▓ŢČF(xiĘĄn)┤Š┴┐����íóÁ˛┬ń╬´┴┐íó╬´ĚNöÁ(shĘ┤)║═Í▓▒╗╔wÂ╚���úČă´╝żťy(cĘĘ)ÂĘ0-5cm═┴îË┴úĆŻ║═0-30cmËđÖC(jĘę)┘|(zhĘČ)��íúÂČ╝żŢć─┴▓ŢÁěúČ├┐─ŕ11ď┬Í┴12ď┬ÍđĐ«Ţć─┴�����úČ├┐đíů^(qĘ▒)▀B└m(xĘ┤)Ě┼─┴5d��úČŢć─┴Í▄Ă┌15d�úČĎ╗─ŕŢć─┴3┤╬íú
7.2.1 ▓ŢÁě║¤└ÝĚ┼─┴╝╝đg(shĘ┤)í¬┤_ÂĘ▀mĎ╦Ě┼─┴ĆŐ(qiĘóng)Â╚
ď┌Ţć─┴¤ÁŻy(tĘ»ng)Íđ�����úČ┤_ÂĘ▀mĎ╦Á─Ě┼─┴ĆŐ(qiĘóng)Â╚╩ăîŹ(shʬ)đđ▓ŢÁěŻí┐Á╣▄└ÝÁ─╗¨ÁA(chĘ│)íú¤─╝żŢć─┴���úČŢdđˇ┴┐×Ú2.67в/haĽr(shʬ)�����úČ▓ŢÁěż▀ËđÎţ┤ˇÁ─Áě╔¤ČF(xiĘĄn)┤Š┴┐�íó╬´ĚNěSŞ╗Â╚║══┴╚└ËđÖC(jĘę)╠╝║Č┴┐���íú°Ţdđˇ┴┐×Ú5.33в/haĽr(shʬ)��úČ▓ŢÁě═┴╚└╦«ĚÍÎţ║├úĘłD1-22úę���íúŇf(shuĘş)├¸▀mÂ╚Ďř░l(fĘí)─┴▓ŢÁ─│ČĐa(bĘ│)âö╔˙ÚL(zhĘúng)úČżS│Í▓ŢÁě┴╝║├Á─╬´ĚNŻY(jiĘŽ)śő(gĘ░u)║══┴╚└Śl╝■��íú┐╔ĎďŇJ(rĘĘn)×Ú▀mÂ╚Ě┼─┴Ëđ└űË┌▓ŢÁě╗ÍĆ═(fĘ┤)����úČ╩ăżSÎo(hĘ┤)▓ŢÁěŻí┐ÁÁ─ÍěϬ═żĆŻíú
┼»╝żĚ┼─┴�����úČ×ęв╚Ňď÷ÍěĎ╗░Ńď┌2.67в/hm2¤┬ÎţŞ▀úĘłD1-23úęíú2002─ŕ��úČÁ┌1ŢćĚ┼
łD1-22¤─╝żŢć─┴îŽ(duĘČ)Í▓▒╗║══┴╚└Á─Ë░ÝĹ
Îóú║═┴╚└ËđÖC(jĘę)╠╝×Ú2004║═2005─ŕÁ─ĂŻż¨ÍÁ��úČĂńËÓ×Ú2002-2005─ŕĂŻż¨ÍÁ

łD1-23▓╗═ČĚ┼─┴ĆŐ(qiĘóng)Â╚Ě┼─┴×ęв╔˙«a(chĘún)┴ŽÎâ╗»
Îóú║¤─╝ż×Ú2002─ŕ-2006─ŕĂŻż¨ÍÁ��úČÂČ╝ż×Ú2002-2005─ŕĂŻż¨ÍÁ
─┴╔§Í┴┐╔Ďď▀_(dĘó)ÁŻ176.7g/d�úČÁ┌2Ţć║═Á┌3ŢćĚ┼─┴úČĎ▓┐╔Ďďď┌120g/dĎď╔¤��íú5.33в/hm2║═8.67в/haÁ─×ęв╚Ňď÷ÍěĚÍäeď┌65í¬167.5g/d║═66.2g/dí¬138.1g/dÍ«Úg��íú▀^(guĘ░)╚ą5─ŕÍđ����úČĚ┼─┴×ęвć╬Í╗╗ţˇwď÷ÍěŰSĚ┼─┴ĆŐ(qiĘóng)Â╚ď÷┤ˇ│╩ŻÁÁ═┌ůäŢ(shĘČ)��íúď┌Ď╗ÂĘŢdđˇ┴┐ĚÂç˙â╚(nĘĘi)�����úČć╬╬╗├ŠĚe▓ŢÁěĚ┼─┴×ęвÁ─ĂŻż¨╚Ňď÷Íě│╩╔¤╔ř┌ůäŢ(shĘČ)�íúϲ?yĘĄn)Ú«?dĘíng)Ě┼─┴ĆŐ(qiĘóng)Â╚│Č▀^(guĘ░)Ď╗ÂĘĚÂç˙║ˇúČË╔Ë┌▓ŢÁě╦ąöí��úČ╝Ďđˇ╔˙«a(chĘún)─▄┴Ž╝▒äí¤┬ŻÁíú
Ď└ô■(jĘ┤)Í▓▒╗┼c═┴╚└╠ěҸ���úČ┤_ÂĘďç˛×(yĘĄn)ů^(qĘ▒)▓ŢÁě╔˙ÚL(zhĘúng)╝żúĘ6ď┬í¬9ď┬úę▀mĎ╦Ţdđˇ┴┐×Ú3.6-4.1вć╬╬╗/ha���íú╝ĎđˇÁ─╔˙«a(chĘún)┴Ž╠ěҸĎ▓┐╔Ďďθ×Ú┤_ÂĘ▀mĎ╦Ě┼─┴ĆŐ(qiĘóng)Â╚Á─Ď└ô■(jĘ┤)úĘłD1-23úęíúިô■(jĘ┤)═┴╚└║═Í▓▒╗╦¨┤_ÂĘÁ─▀mĎ╦Ě┼─┴ĆŐ(qiĘóng)Â╚Űm╚╗ď┌▀@éÇ(gĘĘ)ÍÁ˲ĚÂç˙â╚(nĘĘi)����úČÁź▒╚ÎţŞ▀ÍÁÁ═íú▀@Ě┤Ë││÷▓ŢÁě╔˙«a(chĘún)Íđ�����úČŻŤ(jĘęng)Ł˙(jĘČ)đžĎŠúĘĎďäË(dĘ░ng)╬´╔˙«a(chĘún)×Ú┤˙▒Ýúę┼c╔˙ĹB(tĘĄi)đžĎŠúĘĎďÍ▓▒╗║══┴╚└áţŤr×Ú┤˙▒ÝúęÁ─├ČÂ▄���úČÁźÂ■Ň▀╩ă┐╔Ň{(diĘĄo)║═Á─��íú╔˙«a(chĘún)îŹ(shʬ)█`▀^(guĘ░)│╠Íđ���úČ▀mĎ╦Á─Ě┼─┴ĆŐ(qiĘóng)Â╚đŔϬĎ└ô■(jĘ┤)═┴í¬▓Ţí¬đˇżC║¤╠ěҸ┤_ÂĘíú▒żÝŚ(xiĘĄng)Đđż┐Ďď╔˙ĹB(tĘĄi)đžĎŠ×Ú͸��úČ╝ŠţÖŻŤ(jĘęng)Ł˙(jĘČ)đžĎŠ��úČË├═┴í¬▓Ţí¬đˇ¤Ó╗ąŕP(guĘín)¤Á┤_ÂĘ▀mĎ╦Ţdđˇ┴┐×Ú4.3в/haí¬5.6в/ha�����úČ▓ŢÁěÁ─äË(dĘ░ng)╬´╔˙«a(chĘún)─▄┴Ž×Ú41.9APU/haí¬51.2APU/ha���íú
ÂČ╝żŢć─┴ďç˛×(yĘĄn)▒Ý├¸úČ×ęв╔˙«a(chĘún)┴Ž▓╗═Č│╠Â╚¤┬ŻÁúĘłD1-23úę�����úČĚ┼─┴ďŻŢp�úČ×ęвéÇ(gĘĘ)ˇw╩žÍěÎţ╔┘íúĆ─đˇ«a(chĘún)ĂĚ«a(chĘún)┴┐Á─ŻăÂ╚üÝ(lĘói)Ív����úČÂČ╝ż▓╗▀m║¤Ě┼─┴íúÁź╩ă�úČîŽ(duĘČ)Ë┌đˇ«a(chĘún)ĂĚ┘|(zhĘČ)┴┐╔¤╗˛Ň▀╗¨ÁA(chĘ│)đˇ╚║żS│ÍúČůsż▀ËđĎ╗ÂĘÁ─┐╔đđđď�����íú╚š╣űŻY(jiĘŽ)║¤Đa(bĘ│)´Ľ�����úČ═ŕ╚ź┐╔Ďď▒úÎCżdвˇwÍěď÷╝Ë�úČ╔˙«a(chĘún)╔¤Ď▓ż▀Ëđ┐╔đđđďíú
Áź╩ă����úČĆ─đˇ╚║ďŻÂČÁ─ŻăÂ╚úČ2002─ŕ║═2003─ŕ×ęвď┌45dâ╚(nĘĘi)ˇwÍěĚÍäe¤┬ŻÁ5.1%-8.4%║═2.0%-4.0%���íú╝ĎđˇˇwŤr╔đ┐╔ŻË╩▄���úČ°ăĎŤ](mĘŽi)ËđĐa(bĘ│)´Ľ┘M(fĘĘi)Ë├úČŻŤ(jĘęng)Ł˙(jĘČ)╔¤Ď▓┐╔đđ����íú°ăĎĚ┼─┴▓ŢÁě¤ÓîŽ(duĘČ)Ë┌╬┤Ě┼─┴Á─îŽ(duĘČ)ŇŇśËÁěúČÍ▓▒╗║══┴╚└Ť](mĘŽi)Ëđ´@Í°Îâ╗»úĘp>0.05úę�úČ╔˙ĹB(tĘĄi)╔¤┐╔đđíú▀@ÝŚ(xiĘĄng)Đđż┐�����úČ▀ÇđŔŻY(jiĘŽ)║¤Đa(bĘ│)´Ľ▀M(jĘČn)đđ�úČĎď╠Ż╦¸ÂČ╝ż└űË├╠ý╚╗▓ŢÁěÁ─Îţ╝Đ═żĆŻíú
7.2.2▀mĎ╦Ě┼─┴?xʬ)rÚgÁ─┤_ÂĘ----Ţć─┴▓ŢÁěŻí┐Á╣▄└ÝÁ─í░Úô¤ŮŰpϲÎËĚĘí▒
ިô■(jĘ┤)─┴▓Ţ╔˙ÚL(zhĘúng)äË(dĘ░ng)ĹB(tĘĄi)����úČĎ╗éÇ(gĘĘ)Ţć─┴Í▄Ă┌┐╔ĎďĚÍ×ÚĚ┼─┴Ă┌║═ď┘╔˙ÚL(zhĘúng)Ă┌úĘłD1-24úę���íú▓ŢÁěĚ┼─┴«a(chĘún)╔˙Ď╗éÇ(gĘĘ)Í▓╬´╔˙└Ý╔˙ÚL(zhĘúng)Á─Á═¤ŮúČ▀@╩ă▓ŢÁěżS│Í┐╔│Í└m(xĘ┤)╣Ž─▄║═Żí┐Á╦¨─▄│đ╩▄Á─ÎţÁ═ÚôÍÁ�����úČîŽ(duĘČ)Ŭ(yĘęng)Í°▓ŢÁěÎţÁ═Ş▀Â╚�íó╔wÂ╚╗˛ČF(xiĘĄn)┤Š┴┐Á╚íú─┴▓ŢÁ─ď┘╔˙ÚL(zhĘúng)Ă┌ËÍ┐╔ĎďĚÍ×ÚŇ{(diĘĄo)ŇűĂ┌║═╗ÍĆ═(fĘ┤)Ă┌úĘłD1-24úę����íú─┴▓Ţď┌Ň{(diĘĄo)ŇűĂ┌͸ϬđŮĆ═(fĘ┤)╩▄ôp╔˙└ÝÖC(jĘę)śő(gĘ░u)úČÍěđ┬ĚÍ┼ń┘A▓ě╬´┘|(zhĘČ)���úČ×Ú╣┌îËÍěŻĘ║═ď┘╔˙ÚL(zhĘúng)θ▒ěϬÁ─╔˙└Ý╔˙ÚL(zhĘúng)ť╩(zhĘ│n)éń���íú─┴▓Ţď┌╗ÍĆ═(fĘ┤)Ă┌╔˙└Ý╔˙ÚL(zhĘúng)╣Ž─▄ÍŁu╗ÍĆ═(fĘ┤)ÁŻĚ┼─┴ă░Á─Żí┐Á╦«ĂŻúČ▓ó«a(chĘún)╔˙Ď╗éÇ(gĘĘ)╔˙└Ý╔˙ÚL(zhĘúng)Á─╔¤¤ŮúĘłD1-24úę�����úČ▀@╩ă─┴▓Ţ╔˙└Ý╔˙ÚL(zhĘúng)╗ţäË(dĘ░ng)─▄ë˛╗ÍĆ═(fĘ┤)Á─ÎţŞ▀ÚôÍÁ�úČîŽ(duĘČ)Ŭ(yĘęng)Í°▓ŢÁěÎţ┤ˇČF(xiĘĄn)┤Š┴┐╗˛┘A▓Ţ┴┐Á╚íúď┌Ě┼─┴Ă┌����úČ─┴▓ŢÁ─╔˙└Ý╔˙ÚL(zhĘúng)ÍŞś╦(biĘío)ŻËŻŘË┌Ňř│úÁ─╔˙ÚL(zhĘúng)Á═¤ŮÚôÍÁúČ╚˘└^└m(xĘ┤)Ě┼─┴�úČät▓ŢÁěŰyĎď╗ÍĆ═(fĘ┤)úČϲ┤╦�����úČ╔˙ÚL(zhĘúng)Á═¤ŮÚôÍÁ┐╔θĚ┼─┴ŻY(jiĘŽ)╩°Á─ÍŞś╦(biĘío)�����íú─┴▓Ţď┘╔˙ÚL(zhĘúng)Ă┌����úČ─┴▓Ţ╔˙ÚL(zhĘúng)▀_(dĘó)ÁŻ╔˙ÚL(zhĘúng)Ş▀¤ŮÚôÍÁúČ╚˘▓ŢÁě╚ď▓╗└űË├�úČż═Ľ■(huĘČ)ďý│╔┘Yď┤└╦┘M(fĘĘi)úČ°ăĎ▓Ţ╚║ŻY(jiĘŽ)śő(gĘ░u)Îâë─�úČ╔˙ÚL(zhĘúng)Ş▀¤ŮÚôÍÁ┐╔θ×ÚĚ┼─┴Ú_╩╝Á─ÍŞś╦(biĘío)íú╦¨Ďď�úČ═Ę▀^(guĘ░)ďç˛×(yĘĄn)┤_ÂĘ▀mĎ╦Á─Á═¤ŮÚôÍÁ║═Ş▀¤ŮÚôÍÁúČ┐╔ĎďÍŞîž(dĘúo)▓ŢÁěÁ─Żí┐Á╣▄└ÝúČ▀@ż═╩ă▓ŢÁěŻí┐Á╣▄└ÝÁ─ŰpϲÎËÚô¤ŮĚĘ�����íúîŽ(duĘČ)Ë┌Ě┼─┴▓ŢÁě���úČ─┴▓Ţď┌ø╣┘����íóéÇ(gĘĘ)ˇw�íóĚN╚║║═╚║┬ńÁ╚޸éÇ(gĘĘ)ŻM┐ŚîË┤╬Â╝┤Šď┌â╔éÇ(gĘĘ)íúď┌╠ý╚╗▓ŢÁě���úČË╔Ë┌╝ĎđˇÁ─ô˝╩│đď║═─┴▓Ţ┐╣─┴đďÁ─ĚNÚg▓ţ«É�����úČ─┴▓ŢĚNÍ«ÚgÁ─╔˙└Ý╔˙ÚL(zhĘúng)Îâ╗»▓╗═Č▓Ż����úČŞ¸ÝŚ(xiĘĄng)ÍŞś╦(biĘío)Îâ«É┬╩║▄Ş▀����úČϲ┤╦┤_ÂĘŰpϲÎËÁ─╚║ˇwÚô¤ŮÍÁŞŘż▀îŹ(shʬ)█`âr(jiĘĄ)ÍÁúČ╚š╚║┬ń╔wÂ╚íóÁě╔¤ČF(xiĘĄn)┤Š┴┐�íóÁ˛┬ń╬´ČF(xiĘĄn)┤Š┴┐Á╚úČÂ╝╩ă╚║┬ń╔˙«a(chĘún)┴ŽÁ─▒ÝČF(xiĘĄn)đ╬╩Ż���íú
ď┌¤─╝żĚ┼─┴ÁěúČŞ¨ô■(jĘ┤)Úô¤ŮŰpϲÎËĚĘ�����úČ▓╔Ë├ÂÓĚNÍ▓▒╗ÍŞś╦(biĘío)�����úČ┐╔Ďď┤_ÂĘ▓╗═ČŻÁ╦«╠ěҸÁ──ŕĚŢ���úČĚ┼─┴Áě║╬Ľr(shʬ)Ú_╩╝Ě┼─┴║═ŻY(jiĘŽ)╩°Ě┼─┴úĘłD1-24úę�úČ╩╝ŻK▒ú│ÍĚ┼─┴▓ŢÁěÁ─Żí┐ÁáţĹB(tĘĄi)�íúďôĚŻĚĘď┌ďç˛×(yĘĄn)ů^(qĘ▒)Ě┼─┴¤ÁŻy(tĘ»ng)ÍđÁ├ÁŻ▀\(yĘ┤n)Ë├úČ╚íÁ├┴╝║├đž╣ű�íú
łD1-24 ┼»╝żŢć─┴Ă┌ÚgúČ▓ŢÁě╔˙╬´┴┐┼c╬´ĚNäË(dĘ░ng)ĹB(tĘĄi)
7.3 í░═╦ŞűÁěĚÔËří▒Á─Îď╚╗╗ÍĆ═(fĘ┤)╝╝đg(shĘ┤)
ď┌Đđż┐ů^(qĘ▒)ďO(shĘĘ)Í├ĎĐ╗ÍĆ═(fĘ┤)1─ŕ�����íó2─ŕíó4─ŕ║═7─ŕÁ─═╦ŞűÁě����úČĎď╠ý╚╗▓ŢÁěθ×ÚîŽ(duĘČ)ŇŇúČ▒╚Ţ^Í▓▒╗┼c═┴╚└Á─╗ÍĆ═(fĘ┤)▀M(jĘČn)│╠���íú
ŘS═┴Ş▀ďş═╦ŞűÁě╗ÍĆ═(fĘ┤)▀M(jĘČn)│╠Íđ����úČÍ▓▒╗Ć─╔│┼ţć╬â×(yĘşu)╚║┬ń¤˛Ďď▀_(dĘó)ק└´║˙ÍŽÎË×Úâ×(yĘşu)äŢ(shĘČ)ĚNÁ─╚║┬ńĐŢ╠Š�����íú╬´ĚNÁ─ď÷╝Ë╦┘Â╚����úČ│§Ă┌┐ýúČ║ˇĂ┌┬ř����íú═╦Şű║ˇ╗ÍĆ═(fĘ┤)│§Ă┌Á─đ┬ď÷╬´ĚN͸Ϭ╩ă╔│┼ţ║═░Î▓ŢúČÍđă░Ă┌͸Ϭ╩ăĎŕÉŢ´����úČ╦Řéâ╩ă╚║┬ńÁ─┤╬â×(yĘşu)äŢ(shĘČ)ĚN�����ú╗Íđ║ˇĂ┌đ┬ď÷╬´ĚN͸Ϭ╩ăÂÓ─ŕ╔˙Â╣┐ĂÍ▓╬´�úČÍ╗╩ă╚║┬ńÁ─░Ú╔˙ĚN�����íú╚║┬ńÍđ1─ŕ╔˙║═ÂÓ─ŕ╔˙Í▓╬´Áě╔¤╔˙╬´┴┐ĚÍäeĎď8.8%Á──ŕĂŻż¨╦┘┬╩ťp╔┘╗˛ď÷╝Ë�úČÂ╣┐ĂÍ▓╬´Áě╔¤╔˙╬´┴┐▒╚└ř╔¤╔řúĘłD1-25úę�����íú═╦ŞűÁě╗ÍĆ═(fĘ┤)▀^(guĘ░)│╠Íđ�����úČ╔¤îË═┴╚└Íđ╚║┬ńÁě¤┬╔˙╬´┴┐▒╚└ř┼cÎâ╗»Ě¨Â╚ď÷╝Ë���úČ▒ÝČF(xiĘĄn)│÷ި¤Áť\îË╗»┌ůäŢ(shĘČ)�����íú╔│┼ţĚN╚║║═▀_(dĘó)ק└´║˙ÍŽÎËĚN╚║ÍŕŞ▀���íó╣┌̨Í▒ĆŻ����íóĚÍÍŽöÁ(shĘ┤)║═╔˙Í│ÍŽöÁ(shĘ┤)ŰS╗ÍĆ═(fĘ┤)Ľr(shʬ)Úg│╩¤ÓĚ┤Á─Îâ╗»┌ůäŢ(shĘČ)����íú═╦ŞűÁě╗ÍĆ═(fĘ┤)ĐŢ╠Š1í¬7─ŕ═┴╚└Ň│┴ú║═Ě█┴úťp╔┘úČ╔░┴úď÷╝Ë�úČ7─ŕ║ˇÎâ╗»┌ůäŢ(shĘČ)¤ÓĚ┤íú═╦ŞűÁě0-100cm═┴îË╦«ĚÍ║Č┴┐ď┌╗ÍĆ═(fĘ┤)Ă┌Úg│╩ÍŁu╔¤╔ř┌ůäŢ(shĘČ)���úČÍđĂ┌ď÷̨´@Í°�����íú╚źN║═╦┘đžNď┌╗ÍĆ═(fĘ┤)ă░Ă┌ťp╔┘�����úČ║ˇĂ┌ď÷╝Ë����íúěÎŞţ└űË├¤┬�����úČ═╦ŞűÁě╗ÍĆ═(fĘ┤)▀^(guĘ░)│╠Íđ═┴╚└╚źPíó╦┘đžP║═ËđÖC(jĘę)C│Í└m(xĘ┤)╦ąŻ▀úĘłD1-25úę�íú
łD1-25 ═╦ŞűÁě╗ÍĆ═(fĘ┤)▀M(jĘČn)│╠ÍđÍ▓▒╗┼c═┴╚└╠ěҸ
ިô■(jĘ┤)ĚN╚║íó╚║┬ń║══┴╚└╠ěҸ¤ÓîŽ(duĘČ)Ë┌╠ý╚╗▓ŢÁěÁ─Îâ╗»╦┘Â╚����úČ═╦ŞűÁě╗ÍĆ═(fĘ┤)ÁŻ«ö(dĘíng)ă░ĚÇ(wĘžn)ÂĘÁ─╚║┬ń╦¨đŔĽr(shʬ)ÚgĚÍäe×ÚúČâ×(yĘşu)äŢ(shĘČ)ĚN╚║8í¬9─ŕ�����úČ╚║┬ń9í¬11─ŕ�úČ═┴╚└11í¬12─ŕúĘ▒Ý1-27úę�íúÚL(zhĘúng)Ă┌ŞűθŞ─╔Ă┴╦═┴╚└ŻY(jiĘŽ)śő(gĘ░u)┼cĚ╩┴ŽúČϲ┤╦ď┌═╦ŞűÁěŚëŞűďšĂ┌�����úČ╚║┬ńż▀ËđŢ^Ş▀Á─╔˙╬´┴┐���ú╗ŚëŞűÁěÁ─╗ÍĆ═(fĘ┤)ĐŢ╠Š╦┘Â╚Ď▓▒╚╚║┬ńÁ─ďş╔˙ĐŢ╠Š┐ýÁ├ÂÓ�����úČ▓óăĎŚëŞűÁěĆ─Ď╗Ú_╩╝ż═ż▀ËđŢ^Ş▀Á─╔˙ĹB(tĘĄi)Ě└Îo(hĘ┤)─▄┴Ž�íú╦¨ĎďúČď┌Đđż┐ů^(qĘ▒)˲����úČÂÓöÁ(shĘ┤)ŞűÁěË├ŚëŞűĚÔËřÁ─ĚŻ╩ŻÚ_Ň╣╔˙ĹB(tĘĄi)╗ÍĆ═(fĘ┤)╩ă┐╔đđÁ─íú
▒Ý1-27 ═╦ŞűÁěÁ─╗ÍĆ═(fĘ┤)Ľr(shʬ)Úg
|
|
ÍŞś╦(biĘío) |
╗ÍĆ═(fĘ┤)Ľr(shʬ)Úg úĘ─ŕúę |
|
ĚN╚║ |
╔│┼ţ |
8.4í└0.2 |
|
▀_(dĘó)ק└´║˙ÍŽÎË |
8.9í└0.4 |
|
ĂŻż¨ |
8.7í└0.3 |
|
╚║┬ń |
╚║┬ń¤Ó╦Ăđď |
9.5 |
|
╬´ĚNěSŞ╗Â╚ |
10.4 |
|
Áě╔¤ČF(xiĘĄn)┤Š┴┐ |
12.6 |
|
ÂÓ─ŕ╔˙Í▓╬´/1─ŕ╔˙Í▓╬´Áě╔¤╔˙╬´┴┐▒╚└ř |
8.6 |
|
ĂŻż¨ |
10.3í└0.9 |
|
═┴╚└
|
═┴╚└┴úĆŻúĘŇ│┴ú����íóĚ█┴ú║═╔│┴úúę |
11.4 |
|
═┴╚└Ě╩┴ŽúĘ╚źÁ¬║═╦┘đžÁ¬úę |
11.9 |
|
ĂŻż¨ |
11.6í└0.4 |
8. ═╦╗»íó╔│╗»▓ŢÁěÍ▓▒╗╗ÍĆ═(fĘ┤)╝╝đg(shĘ┤)Đđż┐
┴Í╬¸┐hËđŻŘ10╚f(wĘĄn)╣źÝĽÁ─╠ý╚╗▓ŢÁě�úČË╔Ë┌ÚL(zhĘúng)Ă┌│ČŢd▀^(guĘ░)─┴║═├Ą─┐Ú_ëĘúČ▓ŢÁě═╦╗»����íó╔│╗»╚Ň┌ůç└(yĘón)ÍěúČ═╦╗»���íó╔│╗»▓ŢÁě├ŠĚeŇ╝┐╔└űË├▓ŢÁě├ŠĚe85%Ďď╔¤����íú×Ú┴╦╗ÍĆ═(fĘ┤)▓ŢÁěÍ▓▒╗�����úČ╠ߪ▀▓ŢÁě╔˙«a(chĘún)┴ŽúČ╬ĎéâÚ_Ň╣┴╦═╦╗»����íó╔│╗»╠ý╚╗▓ŢÁěÍ▓▒╗╗ÍĆ═(fĘ┤)╝╝đg(shĘ┤)Đđż┐┼c╩żĚÂ═ĂĆVíúď┌╔ţ╚ŰĐđż┐╚ź┐h▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╩▄ôpÖC(jĘę)└ÝÁ─╗¨ÁA(chĘ│)╔¤�úČ▀\(yĘ┤n)Ë├ç˙ĚÔŻű─┴íóđŢ─┴Á╚Îď╚╗╗ÍĆ═(fĘ┤)║═▀xô˝▀mŬ(yĘęng)đďĆŐ(qiĘóng)Á──┴▓Ţ▀M(jĘČn)đđĐa(bĘ│)▓ąÁ╚╝╝đg(shĘ┤)┤Ű╩ę���úČ╣▓Ş─┴╝╔│╗»�íó═╦╗»�����íóű}ëA╗»▓ŢÁě4100hm2���úČ╩╣Í▓▒╗╔wÂ╚Ć─ďşüÝ(lĘói)▓╗ÎŃ30%╠ߪ▀ÁŻ80%Ďď╔¤úČ─┴▓Ţ«a(chĘún)┴┐Ć─434.69íź750.01kg/hm2╠ߪ▀ÁŻ1464.04íź2850.12kg/hm2���úČÁě¤┬╔˙╬´┴┐Ď▓├¸´@ď÷╝Ë���úČ▓ŢÁěç˙ĚÔ4─ŕ║ˇúČÁě¤┬╔˙╬´┴┐Ë╔ďşüÝ(lĘói)Á─4.41gď÷╝ËÁŻ28.49g(ÚL(zhĘúng)í┴îĺí┴╔ţÂ╚=20cmí┴20cmí┴30cm)�����íú╔│╗»íó═╦╗»▓ŢÁěÍ▓▒╗Á├ÁŻ├¸´@Á─╗ÍĆ═(fĘ┤)�úČ╚║┬ńŻY(jiĘŽ)śő(gĘ░u)░l(fĘí)╔˙´@Í°Îâ╗»úČĆ─°¢ÍĂ┴╦▓ŢÁě═╦╗»Á─ĹB(tĘĄi)äŢ(shĘČ)�����íú
▀xô˝ŢpÂ╚╗˛ÍđÂ╚═╦╗»▓ŢÁě▀M(jĘČn)đđç˙ĚÔŻű─┴�����úČ4─ŕ║ˇ���úČ▓ŢÁěÍ▓▒╗Á├ÁŻ├¸´@Á─╗ÍĆ═(fĘ┤)����úČ╚║┬ńŻY(jiĘŽ)śő(gĘ░u)░l(fĘí)╔˙├¸´@Îâ╗»�úČÍ▓╬´ĚNţÉď÷ÂÓúČË╔ďşüÝ(lĘói)Á─╩«ÄÎĚNď÷╝ËÁŻÄÎ╩«ĚNúĘ▒Ý1-28úę�����íú
▒Ý1-28ííÍ▓╬´╚║┬ńŻY(jiĘŽ)śő(gĘ░u)╠ěđď
|
śËÁě
├ű |
└űË├
ĚŻ╩Ż |
â×(yĘşu) äŢ(shĘČ)
Í▓ ╬´ |
Í▓ ╬´ ĚN ţÉ |
|
┤ˇ╦«▓Ą┴_─┴ł÷(chĘúng)
|
╚ź─ŕĚ┼─┴ |
▓┌Ű[ÎË▓Ţíóßś├ę�����íó
└ăÂż |
└ăÂż�����íó▓Ţ╩|¤Ńíó╗Żq▓Ţíó▓┌Ű[ÎË▓Ţ���íó┤ˇßś├ę�����íóв▓Ţ�íó▓┌╠Kíó▒ÔđţÂ╣�����íó╬»┴ŕ▓╦�íó╠Ă╦╔▓Ţ�����íó╝Ü(xĘČ)╚~╩[íó▓˝║˙���íó┴Đ╚~Ţ´�íóĚ└´L(fĘąng)�íóÁěË▄íó╝Ü(xĘČ)╚~°S╬▓ |
|
░Ű─ŕŻű─┴(ÂČ╝żĚ┼─┴) |
żÇ╚~żŇ����íó
┤ˇßś├ęíó
╝Ô╚~║˙ÍŽÎË |
żÇ╚~żŇ�íó┤ˇßś├ęíó╝Ô╚~║˙ÍŽÎË���íó╗Żq▓Ţ�íó▓Ţ╩|¤Ń����íó└ăÂżíóÁěË▄����íó║ÁűťĂ┐íó░óáľ╠ę╣Ě═Ů╗Ęíó┴Đ╚~Ţ´�����íó▓┌╠K���íó╠Ž▓Ţ�íó▓┌Ű[ÎË▓Ţ����íóăó▓Ţíó╔ńŞ╔°S╬▓�íóŘS▄╬íó´L(fĘąng)ÔĆ▓Ţ���íóÂ■┴Đ╬«┴ŕ▓╦���íó▓˝║˙íó╝ČÂ╣�����íó╔ŻĎ░═ŃÂ╣�����íó▒ÔđţÂ╣�����íó╝Ü(xĘČ)╚~╩[�����íó╩»Í˝���íó┬Ú╗Ęţ^�íó╝Ü(xĘČ)╚~°S╬▓�����íóăě▄┤�����íóđă├ź╬«┴ŕ▓╦�íóŮĄíóđ▒ăoŘS▄╬���íó═▀╦╔�����íóĚ└´L(fĘąng)�íóÂ╠╗Ęßś├ęíó▓Ţ─żÚěáţŘS▄╬���íó▄çŢS▓Ţ���íó┤╠╦ŰŮ╝íóăGŻŠ���íó┼ţÎË▓╦���íó═┴╚řĂ▀íó═ßţ^▓╦����íó└ńŢ´ |
|
đ┬│ăÎËŠé(zhĘĘn) |
╚ź─ŕĚ┼─┴ |
░┘└´¤Ńíó└ńŢ´�����íó╣Ě╬▓▓Ţíó |
▀_(dĘó)ק└´║˙ÍŽÎË�����íó░┘└´¤Ń����íó└ńŢ´�����íó╝ČÂ╣�����íóÁěň\����íó░óáľ╠ę╣Ě╗Ęíó╣Ě╬▓▓Ţ���íó╗ó╬▓▓Ţ |
|
┤˛▓Ţł÷(chĘúng) |
▀_(dĘó)ק└´║˙ÍŽÎË�����íóв▓Ţ����íó
Íđ╚AŰ[ÎË▓Ţ |
▀_(dĘó)ק└´║˙ÍŽÎËíó░┘└´¤Ń���íó└ńŢ´���íó╝ČÂ╣íóÍđ╚AŰ[ÎË▓Ţ�����íóË├ę����íóв▓Ţíó┤ˇ╚~Ň┴���íó▓Ţ─żÚěáţŘS▄╬�íó▓Ţ╩|¤Ń���íó╬«┴ŕ▓╦�íóͬ─Şíó╝Ü(xĘČ)╚~╩[�����íó▓Ţ┬ÚŘS�íó╔ńŞ╔°S╬▓
|
|
Űpż«ÁŕÓl(xiĘíng) |
╚ź─ŕĚ┼─┴ |
Ď╗─ŕ╔˙Űs▓Ţ |
▀_(dĘó)ק└´║˙ÍŽÎËíó└ńŢ´�����íó╝ČÂ╣���íó▓┌╚~ŘS▄╬íó╝Ü(xĘČ)╚~°S╬▓�����íóÁěň\���íóÍ╣Ь˝R╠Ă�����íó┤š▓Ţ╠Ž�����íó╣Ě╬▓▓Ţ |
|
╚ź─ŕŻű─┴ |
в▓Ţ���íó
▒¨▓Ţ���íó
▀_(dĘó)ק└´║˙ÍŽÎË |
в▓Ţíó▒¨▓Ţ���íó▀_(dĘó)ק└´║˙ÍŽÎË�����íó▓┌Ű[ÎË▓Ţ�íó«ő├╝▓Ţ���íó░Î▓Ţ����íó╬«┴ŕ▓╦���íó└ńŢ´�íó╝Ü(xĘČ)╚~°S╬▓íóÁěň\����íó┤š▓Ţ╠Žíó▒ÔđţÂ╣����íó┘ç▓Ţíó╬¸░ě└űüćË├ę�íó░ČŢ´íó└ńŢ´�íóÁě?zĘŽ)ř╣¤íóěi├ź▓╦�íó╗ó╬▓▓Ţ���íó╣Ě╬▓▓Ţ |
▓ŢîËŞ▀Â╚Ë╔ďşüÝ(lĘói)Á─▓╗ÎŃ10cm╠ߪ▀ÁŻ20íź35cm �����úČâ×(yĘşu)┴╝Â╣┐Ă─┴▓Ţ║═║╠▒ż┐Ă─┴▓Ţ├¸´@ď÷ÂÓ�����ú╗▓ŢÁě«a(chĘún)▓Ţ┴┐Á├ÁŻ´@Í°Á─╠ߪ▀���úČĂńÍđÂ╣┐Ă║═║╠▒ż─┴▓Ţ╦¨Ň╝▒╚└řď÷╝Ë(▒Ý1-29)����íú
▒Ý1-29╠ý╚╗▓ŢÁěÁě╔¤╔˙╬´┴┐úĘ kg/hm2úę
|
śËÁě├ű |
└űË├ĚŻ╩Ż |
┐é╔˙╬´┴┐ |
Â╣┐Ă─┴▓Ţ |
║╠▒ż┐Ă
─┴▓Ţ |
ŰsţÉ▓Ţ |
|
┤ˇ╦«▓Ą┴_─┴ł÷(chĘúng)
┤ˇ╦«▓Ą┴_─┴ł÷(chĘúng)
đ┬│ăÎËŠé(zhĘĘn)
đ┬│ăÎËŠé(zhĘĘn)
Űpż«ÁŕÓl(xiĘíng)
Űpż«ÁŕÓl(xiĘíng)
┼d┬íăfÓl(xiĘíng)
┼d┬íăfÓl(xiĘíng) |
╚ź─ŕĚ┼─┴
░Ű─ŕŻű─┴
╚ź─ŕĚ┼─┴
┤˛▓Ţł÷(chĘúng)
╚ź─ŕĚ┼─┴
╚ź─ŕŻű─┴
´w▓ąĚÔËř┤˛▓Ţ
ť\ŞűĐa(bĘ│)▓ą┤˛▓Ţ |
426.21
1874.24
436.69
902.25
119.66
687.58
1112.86
1464.04 |
138.97
53.65
214.61
155.18
914.96
640.32 |
200.20
321.06
294.50
69.06
41.28
236.17
179.89
427.52 |
226.01
1414.21
88.54
618.61
74.38
296.23
18.01
396.20 |
╔│╗»�����íó═╦╗»▓ŢÁěď┌Żű─┴�����íóĚÔËřÁ─▀^(guĘ░)│╠Íđ����úČ│řÁě╔¤▓┐ĚÍÍ▓▒╗Á├ÁŻ╗ÍĆ═(fĘ┤)═ÔúČÁě¤┬╔˙╬´┴┐Ď▓░l(fĘí)╔˙┴╦├¸´@Á─Îâ╗»(▒Ý1-30)�íú
▒Ý1-30 ╠ý╚╗▓ŢÁěÁě¤┬╔˙╬´┴┐ (gíócm)
|
śËÁě├ű |
└űË├ĚŻ╩Ż |
┐骨┴┐ |
ިîË |
|
0íź10 |
10íź20 |
20íź30 |
|
┤ˇ╦«▓Ą┴_─┴ł÷(chĘúng)
┤ˇ╦«▓Ą┴_─┴ł÷(chĘúng)
đ┬│ăÎËŠé(zhĘĘn)
đ┬│ăÎËŠé(zhĘĘn)
Űpż«ÁŕÓl(xiĘíng)
Űpż«ÁŕÓl(xiĘíng) |
╚ź─ŕĚ┼─┴
░Ű─ŕŻű─┴
╚ź─ŕĚ┼─┴
┤˛▓Ţł÷(chĘúng)
╚ź─ŕĚ┼─┴
╚ź─ŕŻű─┴ |
19.84
32.15
4.41
28.49
1.99
12.74 |
17.21
25.45
3.95
25.75
0.82
9.98 |
2.03
3.95
0.24
1.67
0.74
1.85 |
0.60
2.75
0.22
1.07
0.43
0.91 |
|
▀xô˝╔wÂ╚Á═Ë┌30%Á─ÍěÂ╚═╦╗»▓ŢÁě║═╔│╗»▓ŢÁě▀M(jĘČn)đđť\ŞűĐa(bĘ│)▓ą║═╚╦╣Ą─úöM´w▓ą���úČ╚íÁ├┴╦Ţ^║├Á─đž╣ű�����úČĐa(bĘ│)▓ą▓ŢĚN▀xô˝▀mŬ(yĘęng)đďĆŐ(qiĘóng)���íó─═┤ÍĚ┼╣▄└ÝÁ─╝Ô╚~║˙ÍŽÎË���íó▀_(dĘó)ק└´║˙ÍŽÎËíó▒Ô╦Ű▒¨▓Ţ║═├╔╣┼▒¨▓Ţ▀M(jĘČn)đđ═╦╗»▓ŢÁěť\ŞűĐa(bĘ│)▓ą����úČ«a(chĘún)▓Ţ┴┐┐╔▀_(dĘó)1464.04kg/hm2úČĂńÍđ�úČÂ╣┐Ă─┴▓Ţ║═║╠▒ż┐Ă─┴«a(chĘún)┴┐ĚÍäe▀_(dĘó)ÁŻ640.32 kg/hm2║═427.52 kg/hm2ú╗╔│╗»▓ŢÁě╚╦╣Ą─úöM´w▓ą«a(chĘún)▓Ţ┴┐┐╔▀_(dĘó)1112.86 kg/hm2�úČĂńÍđÂ╣┐Ă─┴▓Ţ║═║╠▒ż┐Ă─┴▓Ţ«a(chĘún)┴┐ĚÍäe▀_(dĘó)ÁŻ914.96 kg/hm2║═179.89 kg/hm2íúť\ŞűĐa(bĘ│)▓ąîŽ(duĘČ)ďş╩╝Í▓▒╗Á─ĂĂë─đďŢ^┤ˇ�����úČŞ─┴╝║ˇ1íź2─ŕ▓ŢÁě╔˙«a(chĘún)┴ŽŢ^Á═�����úČÁźÁ┌3─ŕĎď║ˇ«a(chĘún)▓Ţ┴┐äíď÷�����úČ×ÚĚ└Í╣═┴Áě╔│╗»ĂĎŐ�����úČďô╝╝đg(shĘ┤)ď┌▓╔Ë├Ľr(shʬ)Ŭ(yĘęng)╔¸Íě���íú╚╦╣Ą─úöM´w▓ąîŽ(duĘČ)▓ŢÁěďş╩╝Í▓▒╗▓╗Ľ■(huĘČ)ďý│╔ĂĂë─����úČ▀mĎ╦ď┌╔│╗»▓ŢÁě╔¤îŹ(shʬ)╩ę����úČ▀@ĚN╝╝đg(shĘ┤)▓┘θ║ć(jiĘún)ć╬úČ▒ŃË┌╚║▒ŐŇĂ╬Ň����úČ╩ăŞ─┴╝╔│╗»▓ŢÁěđđÍ«ËđđžÁ─║├╝╝đg(shĘ┤)íú
═Ę▀^(guĘ░)╠ý╚╗▓ŢÁěÍ▓▒╗╗ÍĆ═(fĘ┤)╝╝đg(shĘ┤)Đđż┐����úČ╚íÁ├┴╦╚š¤┬ŻY(jiĘŽ)╣űú║ó┘ď┌ŢpÂ╚╗˛ÍđÂ╚═╦╗»▓ŢÁě╔¤úČ▓╔Ë├Żű─┴ĚÔËř╝╝đg(shĘ┤)�úČ┐╔╚íÁ├Ţ^║├Á─đž╣űúČŻŤ(jĘęng)▀^(guĘ░)4─ŕÁ─Żű─┴ĚÔËř�����úČ═╦╗»íó╔│╗»▓ŢÁěÍ▓▒╗Á├ÁŻ├¸´@╗ÍĆ═(fĘ┤)����úČ╚║┬ń░l(fĘí)Ň╣▀M(jĘČn)Ň╣ĐŢ╠Šú╗ó┌ď┌═╦╗»▓ŢÁěĐŢÎâ▀^(guĘ░)│╠Íđ�����úČ╚║┬ńÍđÁ─â×(yĘşu)äŢ(shĘČ)ĚN╚║┤Šď┌Í°ůf(xiĘŽ)═ČÎâ╗»�����ú╗ó█┼c═╦╗»▓ŢÁě¤Ó▒╚�����úČ╔│╗»▓ŢÁěÎď╚╗ĚÔËř╦┘Â╚Ţ^┬ř���úČ╠ěäe╩ăÁě¤┬▓┐ĚÍ����ú╗ó▄╚╦╣Ą─úöM´w▓ą╝╝đg(shĘ┤)▓┘θ║ć(jiĘún)ć╬����úČď┌ÍěÂ╚═╦╗»Á─░Ű┴¸äË(dĘ░ng)╔│Áě▓╔Ë├ďô╝╝đg(shĘ┤)úČ─▄ĐŞ╦┘ď÷╝ËÍ▓▒╗�úČđ╬│╔Ďď´w▓ąâ×(yĘşu)┴╝─┴▓Ţ×Úâ×(yĘşu)äŢ(shĘČ)Á─Í▓╬´╚║┬ńúČď┌ĚÔËřŚl╝■¤┬â╔─ŕ╝┤┐╔╚íÁ├´@Í°│╔đž����íú
9. ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁďu(pʬng)╣└╝░ĂńäË(dĘ░ng)ĹB(tĘĄi)─úöM
▒żĐđż┐╩ăď┌îŽ(duĘČ)Áńđ═ů^(qĘ▒)˲Ň{(diĘĄo)▓ÚĐđż┐Á─╗¨ÁA(chĘ│)╔¤úČ▀\(yĘ┤n)Ë├╔˙ĹB(tĘĄi)ŻŤ(jĘęng)Ł˙(jĘČ)îW(xuĘŽ)����íóđˇ─┴îW(xuĘŽ)íó▓ŢśI(yĘĘ)┐ĂîW(xuĘŽ)Á╚¤ÓŕP(guĘín)îW(xuĘŽ)┐ĂÁ─ͬÎR(shʬ)���úČ▓╔Ë├ĎÄ(guĘę)ĚÂĐđż┐┼cîŹ(shʬ)ÎCĐđż┐¤ÓŻY(jiĘŽ)║¤�����úČÂĘđďĚÍ╬÷┼cÂĘ┴┐ĚÍ╬÷¤ÓŻY(jiĘŽ)║¤Á─ĚŻĚĘ�úČŻY(jiĘŽ)║¤ňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─îŹ(shʬ)ŰH����úČîŽ(duĘČ)ňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─âr(jiĘĄ)ÍÁ╝░Ăń╣▄└Ý▓▀┬ď▀M(jĘČn)đđ┴╦Đđż┐úČŇ╣╩ż┴╦Đđż┐┐˛╝▄Á─└ÝŇôâr(jiĘĄ)ÍÁ║═Ŭ(yĘęng)Ë├âr(jiĘĄ)ÍÁ�úČ╠ŻËĹ┴╦╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁďu(pʬng)╣└Á─└ÝŇô┼c╝╝đg(shĘ┤)�úČ×Ú▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╣▄└Ý����íó╗ÍĆ═(fĘ┤)┼c▒úÎo(hĘ┤)Á─┐ĂîW(xuĘŽ)ŤQ▓▀╠ß╣ę┴╦└ÝŇôĎ└ô■(jĘ┤)íú
9.1. ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁÁ──▄ÍÁďu(pʬng)╣└┐˛╝▄Đđż┐
ßśîŽ(duĘČ)─┐ă░╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁďu(pʬng)╣└┐˛╝▄┤Šď┌Á─͸Ϭćľ(wĘĘn)ţ}ú║1úę���íóâr(jiĘĄ)ÍÁďu(pʬng)╣└Á─└ÝŇô╔đ╠Äď┌╠Ż╦¸▀^(guĘ░)│╠Íđ���úČ╬┤îóĂń╝{╚Ű═Â┘YĚÍ╬÷┼cŤQ▓▀▀^(guĘ░)│╠ú╗2úę�íóĚÍţÉĆ═(fĘ┤)ŰsúČýoĹB(tĘĄi)ĚÍ╬÷ÂÓ���úČäË(dĘ░ng)ĹB(tĘĄi)ĚÍ╬÷╔┘�úČÂĘâr(jiĘĄ)ÄžËđ├¸´@Á─͸Ë^╔ź▓╩����íúŬ(yĘęng)Ë├─▄ÍÁ└ÝŇô┼c╔˙ĹB(tĘĄi)îW(xuĘŽ)íóŻŤ(jĘęng)Ł˙(jĘČ)îW(xuĘŽ)└ÝŇôËđÖC(jĘę)ŻY(jiĘŽ)║¤���úČ╠ß│÷┴╦▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁÁ──▄ÍÁďu(pʬng)╣└┐˛╝▄úĘłD1-26úę����íú
9.1.1╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─͸Ϭâ╚(nĘĘi)╚Ţ
1)╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)┴óÁěĂŻ┼_(tĘói)Á─âr(jiĘĄ)ÍÁú╗2)╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)┘A┤Š╦«╝░ď¬╦ěÁ─âr(jiĘĄ)ÍÁ���ú╗3)═┴╚└─Ó╠┐╝░ËđÖC(jĘę)┘|(zhĘČ)âr(jiĘĄ)ÍÁú╗4) ╗ţÁ─╔˙╬´┴┐Á─âr(jiĘĄ)ÍÁ����ú╗5)╠ěËđ╬´ĚNÁ─╗¨Ď˛đ┼¤óâr(jiĘĄ)ÍÁíú╚š╣ű─│ĚN╬´ĚN▓óĚă▒żÁě¬Ü(dĘ▓)Ëđ�����úČ┐╔ިô■(jĘ┤)ĂńŇ╝╚źă˛ĚÍ▓╝├ŠĚeÁ─▒╚└ř▀M(jĘČn)đđ┐´╦Ń�����íú
9.1.2 ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╣Ž─▄
╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ě■äŇ(wĘ┤)╣Ž─▄Á─͸Ϭâ╚(nĘĘi)╚ŢËđ╔˙ĹB(tĘĄi)╣Ž─▄���íóŻŤ(jĘęng)Ł˙(jĘČ)╣Ž─▄║═╔šĽ■(huĘČ)╣Ž─▄�����íú
9.1.2.1 ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╔˙ĹB(tĘĄi)╣Ž─▄
▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╩ăĎ╗éÇ(gĘĘ)Ë╔ÜÔ║˛����íóÁěđ╬�íó═┴╚└íó╚Ň╣Ô�����íó┐ŇÜÔíó╦«ĚÍ�����íóÁě¤┬┘Yď┤���íóÍ▓╬´�íó
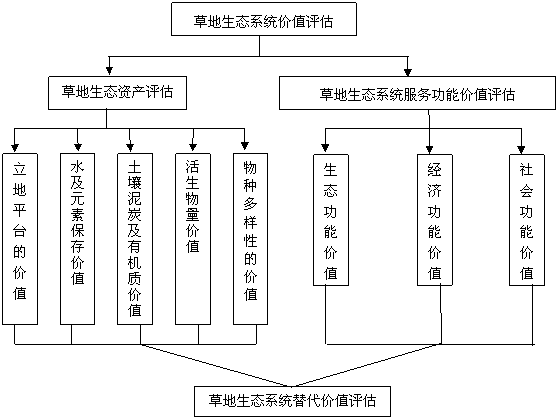
łD1-26 ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁďu(pʬng)╣└┐˛╝▄╝░ĂńÁ─ËđÖC(jĘę)śő(gĘ░u)│╔
äË(dĘ░ng)╬´Ďď╝░╬ó╔˙╬´Á╚ŻM│╔Á─╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)���úČĂń╔˙ĹB(tĘĄi)Ě■äŇ(wĘ┤)╣Ž─▄╩ă▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)┼c╔˙ĹB(tĘĄi)▀^(guĘ░)│╠╦¨đ╬│╔╝░╦¨żS│ÍÁ─╚╦ţÉ┘çĎď╔˙┤ŠÁ─Îď╚╗şh(huĘón)ż│Śl╝■║═đžË├���íú͸Ϭ▒ÝČF(xiĘĄn)╩ăú║Ň{(diĘĄo)╣Ł(jiĘŽ)ÜÔ║˛íóŞ─┴╝═┴╚└�íó▒ú│Í╦«═┴íóĚ└´L(fĘąng)╣╠╔│����íó║şB(yĘúng)╦«ď┤����íó├└╗»şh(huĘón)ż│�íóâ˘╗»┐ŇÜÔíóĚ└Í╣ďŰϢÁ╚ÂÓĚN╔˙ĹB(tĘĄi)Ě■äŇ(wĘ┤)╣Ž─▄���íú═Ę▀^(guĘ░)║¤└ÝÁě░l(fĘí)ô]▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╔˙ĹB(tĘĄi)╣Ž─▄úČ─▄╩╣╚╦ţÉ╔˙╗ţď┌Ď╗éÇ(gĘĘ)ăňŁŹâ×(yĘşu)├└Á─╔˙╗ţşh(huĘón)ż│║═╔˙ĹB(tĘĄi)şh(huĘón)ż│Í«Íđ����úČŢ^║├ÁěĚ└Í╬޸ĚN╬█╚żîŽ(duĘČ)╚╦ţÉÁ─┐╔─▄Ş╔ö_║═╬ú║ŽúČĎď╝░╚╦┐┌Á─Ňř│úď┘╔˙«a(chĘún)║═޸ĚN«a(chĘún)śI(yĘĘ)Á─║¤└Ýď┘╔˙«a(chĘún)�íú
9.1.2.2 ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─ŻŤ(jĘęng)Ł˙(jĘČ)╣Ž─▄
▓ŢÁěÁ─ŻŤ(jĘęng)Ł˙(jĘČ)╣Ž─▄╩ă▓ŢÁ쯍(jĘęng)Ł˙(jĘČ)¤ÁŻy(tĘ»ng)ď┌╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)▀M(jĘČn)đđ╬´┘|(zhĘČ)Đşşh(huĘón)íó─▄┴┐ŮD(zhuĘún)ôQ║═đ┼¤óé¸▀fÁ─═ČĽr(shʬ)�����úČŞ¸ţÉŻŤ(jĘęng)Ł˙(jĘČ)Ϭ╦ěÁ─═Â╚Ű║═«a(chĘún)│÷đ╬│╔┴╦ŁMÎŃ╚╦ţÉ▓╗═ČđŔăˇÁ─޸ĚNËđđ╬║═čo(wĘ▓)đ╬Á─ÍđÚg«a(chĘún)ĂĚ║═ÎţŻK«a(chĘún)ĂĚ���úČď┘═Ę▀^(guĘ░)Ëđđ╬║═Ű[đ╬╩đł÷(chĘúng)Á─Ż╗ôQ�úČ╠ß╣ęŻo▒żÁěů^(qĘ▒)╔˙«a(chĘún)¤ű┘M(fĘĘi)�����íóżË├˝¤ű┘M(fĘĘi)║═ŻŤ(jĘęng)▀^(guĘ░)ç°(guĘ«)â╚(nĘĘi)═Ô┘Q(mĘĄo)ĎÎŁMÎŃç°(guĘ«)â╚(nĘĘi)═Ô╩đł÷(chĘúng)đŔăˇÁ─ÍT╣Ž─▄Á─┐éĚQíú░Ř└Ę▓ŢÁě«a(chĘún)ĂĚ▒ż╔ÝÁ─ŻŤ(jĘęng)Ł˙(jĘČ)╣Ž─▄�����úČ▀ÇËđŞ─╔Ăşh(huĘón)ż│���íó├└╗»╔šů^(qĘ▒)┤┘▀M(jĘČn)Ăń╦ŘđđśI(yĘĘ)����úČ╚šđˇ─┴śI(yĘĘ)�����íó┬├Ë╬śI(yĘĘ)║═Ě┐Áě«a(chĘún)śI(yĘĘ)ď÷ÍÁÁ─╣Ž─▄�����íú
═Ę▀^(guĘ░)░l(fĘí)ô]▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─ŻŤ(jĘęng)Ł˙(jĘČ)╣Ž─▄���úČď┌Îď╚╗╬´┘|(zhĘČ)Đşşh(huĘón)Íđđ╬│╔┴╦╬´┘|(zhĘČ)┴¸����íó─▄┴┐┴¸íóđ┼¤ó┴¸Á─═ČĽr(shʬ)�����úČĎ▓đ╬│╔┴╦ŻŤ(jĘęng)Ł˙(jĘČ)┴¸����íú╝┤Ď╗ĚŻ├Šđ╬│╔┴╦޸ĚN─▄ŁMÎŃ╚╦ţÉđŔăˇÁ─ŻŤ(jĘęng)Ł˙(jĘČ)╬´┘|(zhĘČ)╝┤╩╣Ë├âr(jiĘĄ)ÍÁúČθ×Ú▀@▓┐ĚÍŻŤ(jĘęng)Ł˙(jĘČ)«a(chĘún)ĂĚÁ─╬´┘|(zhĘČ)│đô˙(dĘín)Ň▀���ú╗┴ÝĎ╗ĚŻ├Š����úČď┌▀@▓┐ĚÍŻŤ(jĘęng)Ł˙(jĘČ)«a(chĘún)ĂĚÁ─═Â╚Ű«a(chĘún)│÷▀^(guĘ░)│╠ÍđËÍ▓╗öÓÁěŮD(zhuĘún)ĎĂâr(jiĘĄ)ÍÁ�����íóäô(chuĘĄng)ďýâr(jiĘĄ)ÍÁ║═ď÷ÍÁâr(jiĘĄ)ÍÁ���úČŻŤ(jĘęng)▀^(guĘ░)Ż╗ôQíóĚÍ┼ń�úČÎţŻK▀M(jĘČn)╚ۤű┘M(fĘĘi)úČĎďîŹ(shʬ)ČF(xiĘĄn)Ăńâr(jiĘĄ)ÍÁ���íúĆ─°Ď▓╩╣Îď╚╗─▄┴┐┴¸┼cŻŤ(jĘęng)Ł˙(jĘČ)─▄┴┐┴¸▓ó┤Š����úČ╩╣╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)â╚(nĘĘi)Á─Îď╚╗đ┼¤ó┴¸┼cŻŤ(jĘęng)Ł˙(jĘČ)¤ÁŻy(tĘ»ng)Á─╔˙«a(chĘún)đ┼¤óíó╩đł÷(chĘúng)đ┼¤ó�íó¤ű┘M(fĘĘi)đ┼¤ó▓ó┤Šú╗▓ó╩╣▓ŢÁě╔╠ĂĚ║═▓ŢÁěĚ■äŇ(wĘ┤)ď┌╔˙«a(chĘún)║═┴¸═ĘÍđîŹ(shʬ)ČF(xiĘĄn)âr(jiĘĄ)ÍÁď÷ÍÁ���íú
9.1.2.3 ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╔šĽ■(huĘČ)╣Ž─▄
ď┌▓ŢÁěÍ▓▒╗Á─▒ŐÂÓ╣Ž─▄Íđ�úČ├¸´@▒╗╚╦éâîŹ(shʬ)ŰH╩╣Ë├Á─╣Ž─▄╩ăŁMÎŃżË├˝Ő╩śĚ(lĘĘ)�íó╬─╗»Ż╠Ëříó┐ĂîW(xuĘŽ)ĂŇ╝░Á╚żź╔˝╔˙╗ţÁ─đŔϬ�íúŰSÍ°╔šĽ■(huĘČ)┼c╚╦ţÉÁ─▀M(jĘČn)▓Ż║═░l(fĘí)Ň╣úČżË├˝îŽ(duĘČ)╔˙╗ţşh(huĘón)ż│Á─đŔăˇď┌▓╗öÓ╠ߪ▀���úČđŢÚeÁ─ĎÔÎR(shʬ)ď┌▓╗öÓ╝ËĆŐ(qiĘóng)����í�����ú┐╔ĎďŇf(shuĘş)����úČ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ĎĐŻŤ(jĘęng)│╔×Ú╚╦éâ╔˙╗ţÍđÁ─Ď╗éÇ(gĘĘ)ÍěϬ│╔ĚÍ�úČż▀ËđÍěϬÁ─╔šĽ■(huĘČ)╣Ž─▄�íúĂń╔šĽ■(huĘČ)╣Ž─▄͸Ϭ▒ÝČF(xiĘĄn)×Úú║ŁMÎŃ╔šĽ■(huĘČ)Ş¸ĚŻ├ŠđŔăˇúČ┤┘▀M(jĘČn)żź╔˝╬─├¸ŻĘďO(shĘĘ)�ú╗Ş─╔Ă╔˙«a(chĘún)╔˙╗ţşh(huĘón)ż│úČď÷▀M(jĘČn)╚╦éâŻí┐Á�����ú╗Ş─╔Ă═Â┘Yşh(huĘón)ż│�úČ╬ŘĎřŻĘďO(shĘĘ)┘YŻú╗╠ß╣ę┴╝║├Á─đޤóşh(huĘón)ż│�����úȿƯÔ╚╦éâđ─└Ýë║┴ŽÁ╚�íú
═Ę▀^(guĘ░)╔¤╩÷îŽ(duĘČ)▓ŢÁěÍ▓▒╗¤ÁŻy(tĘ»ng)Á─╠ěŘc(diĘún)╝░ĂńŻY(jiĘŽ)śő(gĘ░u)Á─ĚÍ╬÷�����úČ┐╔Ďď░l(fĘí)ČF(xiĘĄn)���úČ▓ŢÁě¤ÁŻy(tĘ»ng)╩ăĎ╗éÇ(gĘĘ)ÂÓď¬Á─Ć═(fĘ┤)║¤¤ÁŻy(tĘ»ng)�úČϲ┤╦úČ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)▒ě╚╗ż▀Ëđ╔˙ĹB(tĘĄi)�íóŻŤ(jĘęng)Ł˙(jĘČ)┼c╔šĽ■(huĘČ)╚źĚŻ╬╗Á─Ě■äŇ(wĘ┤)╣Ž─▄íú
▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╩ăĎ╗éÇ(gĘĘ)Ú_Ě┼Á─¤ÁŻy(tĘ»ng)����úČ┼c═ÔŻš┤Šď┌Í°─▄┴┐╬´┘|(zhĘČ)Á─Ż╗ôQúČϲ°Ăńż▀ËđÎď╬Ďď÷ÍÁÁ─╣Ž─▄���íú×Ú┴╦┴╦ŻÔ╣Ž─▄Á─äË(dĘ░ng)ĹB(tĘĄi)���úČ╬Ďéâ┐╔Ďď▀xô˝Í¸Ď¬Á─ÍŞś╦(biĘío)▀M(jĘČn)đđäË(dĘ░ng)ĹB(tĘĄi)─úöMíú╚šîŽ(duĘČ)Ë┌▓ŢÁěÍ▓▒╗�����úČ╬Ďéâ┐╔Ďď▀xô˝╗ţ╔˙╬´┴┐���íóâ˘╔˙«a(chĘún)┴Ž����íó╬Ř╠╝Ě┼Ч╣Ž─▄Á╚ŕP(guĘín)ŠIÍŞś╦(biĘío)îŽ(duĘČ)╔˙ĹB(tĘĄi)┘Y«a(chĘún)�íóĚ■äŇ(wĘ┤)╣Ž─▄Á─ŕP(guĘín)ŠIÍŞś╦(biĘío)▀M(jĘČn)đđżC║¤ĚÍ╬÷íú╚˘Ď¬┴╦ŻÔŞŘÂÓ╣Ž─▄Á─äË(dĘ░ng)ĹB(tĘĄi)����úČ┐╔ď÷╝ËĚÍ╩Ď─úđ═Á─â╚(nĘĘi)╚Ţ���íú
9.2 ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░Ě■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁżC║¤ďu(pʬng)╣└▒Ý
ިô■(jĘ┤)─▄ÍÁĚÍ╬÷ĚŻĚĘ║═▓ŻˇE(łD1-27)îŽ(duĘČ)├┐Ď╗ĚN┘Yď┤Á──▄┴┐íó╬´┘|(zhĘČ)▀M(jĘČn)đđĚÍ╬÷�����úČ╚íÁ├Ď╗¤Á┴đ╗¨▒żöÁ(shĘ┤)ô■(jĘ┤)�úČď┌┤╦╗¨ÁA(chĘ│)╔¤żÄÍĂ╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░ĂńĚ■äŇ(wĘ┤)╣Ž─▄żC║¤ďu(pʬng)╣└▒Ý(▒Ý1-31║═▒Ý1-32)íú
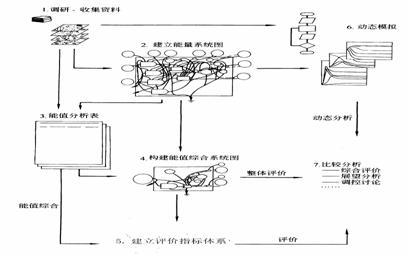
łD1-27─▄ÍÁĚÍ╬÷▓ŻˇE(Ě┬╦{(lĘón)╩óĚ╝�úČ2002;Odum,1996)
▒Ý1-31 ─▄ÍÁĚÍ╬÷▒Ý
|
┘Yď┤ţÉäe |
┘Yď┤┴¸äË(dĘ░ng)┴┐úĘć╬╬╗/─ŕúę |
╠źŕľ(yĘóng)─▄ÍÁŮD(zhuĘún)ôQ┬╩úĘsej/Júę |
╠źŕľ(yĘóng)─▄ÍÁúĘsejúę |
║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ
úĘEM$úę |
|
1.▒żÁě┘Yď┤
1úę
2úę
3úę
ú«ú«ú«
2.Ţö╚Ű┘Yď┤
ú«ú«ú«
3.Ţö│÷┘Yď┤
ú«ú«ú« |
|
|
|
|
▒Ý1-32 ▓ŢÁě╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░ĂńĚ■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁżC║¤ďu(pʬng)╣└▒Ý
|
┘Yď┤ţÉäe |
┘Yď┤┴¸äË(dĘ░ng)┴┐ |
ć╬╬╗ |
╠źŕľ(yĘóng)─▄ÍÁŮD(zhuĘún)ôQ┬╩ |
╠źŕľ(yĘóng)─▄ÍÁúĘsejúę |
║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁúĘEM$úę |
|
1.Ě■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁ
1úę Ң░l(fĘí)
2úę GPP
3úę NPP
1úę ╠╝ůR
2úę ßîĚ┼ЧÜÔ
3úę «a(chĘún)ĂĚ
4úę Ő╩śĚ(lĘĘ)
ú«ú«ú«
2.╔˙ĹB(tĘĄi)┘Y«a(chĘún)âr(jiĘĄ)ÍÁ
1úę┴óÁěĂŻ┼_(tĘói)âr(jiĘĄ)ÍÁ
2úę╗ţÁ─╔˙╬´┴┐
3úę╬´ĚN╗¨Ď˛
4úę─Ó╠┐╝░ËđÖC(jĘę)┘|(zhĘČ)
ú«ú«ú«
3.╠Š┤˙âr(jiĘĄ)ÍÁ
1úę▓┐ĚÍ╠Š┤˙âr(jiĘĄ)ÍÁúĘ╚š▒ÝîË╔˙╬´┴┐╠Š┤˙âr(jiĘĄ)ÍÁúę
2úę═ŕ╚ź╠Š┤˙âr(jiĘĄ)ÍÁ |
|
|
|
|
|
9.3 ňa┴Í╣¨└Ňв▓ŢśËĚŻ▓ŢÁě╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░╣Ž─▄âr(jiĘĄ)ÍÁÁ──▄ÍÁďu(pʬng)╣└
×Ú┴╦╠ŻËĹňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─âr(jiĘĄ)ÍÁúČ▀xô˝╬╗Ë┌ďôů^(qĘ▒)ż│â╚(nĘĘi)Á─Íđç°(guĘ«)┐ĂîW(xuĘŽ)ď║â╚(nĘĘi)├╔▓Ţďş╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ÂĘ╬╗ŇżÁ─śËÁě┘Y┴¤×Ú╗¨ÁA(chĘ│)�����úČďçłD═Ę▀^(guĘ░)░Ş└řĐđż┐�úČŇ╣╩ż▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁďu(pʬng)╣└Á─đ┬ĚŻĚĘíú
▒żĐđż┐Ďďв▓ŢśËÁě┘Y┴¤×Ú└ř▀M(jĘČn)đđ┴╦Đđż┐���íúв▓ŢśËÁěŻĘË┌1980─ŕúČ├ŠĚe400m*600m���íú
9.3.1 ć╬╬╗├ŠĚe─▄ÍÁďu(pʬng)╣└
â╚(nĘĘi)├╔╣┼ňa┴Í╣¨└ŇÁńđ═▓ŢÁěв▓Ţ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ć╬╬╗├ŠĚe─▄ÍÁďu(pʬng)╣└ŻY(jiĘŽ)╣ű┴đË┌▒Ý1-33Íđ���úČ▓óîŽ(duĘČ)ËđŕP(guĘín)˛î(qĘ▒)äË(dĘ░ng)┴Ž����íóşh(huĘón)ż│Ě■äŇ(wĘ┤)╝░╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁĎď─▄ÍÁď¬(em¥)×Úć╬╬╗▀M(jĘČn)đđ┴╦╣└╦Ń�����íú
|
▒Ý1-33 |
|
|
đ˛╠ľ(hĘĄo) |
ÝŚ(xiĘĄng)─┐ |
öÁ(shĘ┤)ô■(jĘ┤)╝░ |
ć╬╬╗ |
─▄ÍÁŮD(zhuĘún)ôQ┬╩ |
╠źŕľ(yĘóng)─▄ÍÁ |
║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ |
|
|
|
|
|
(sej/ć╬╬╗) |
(sej/a) |
(ď¬╚╦├˝Ä┼) |
|
|
˛î(qĘ▒)äË(dĘ░ng)┴Žú║─▄┴┐üÝ(lĘói)ď┤ |
|
|
|
|
|
|
1 |
╠źŕľ(yĘóng)ŢŚ╔ń |
4.43E+13 |
J/hm2/a |
1 |
4.43E+13 |
88.60 |
|
2 |
´L(fĘąng) |
3.10E+10 |
J/hm2/a |
1496 |
4.64E+13 |
92.75 |
|
3 |
Ëŕ╦«ú║╗»îW(xuĘŽ)─▄ |
1.73E+10 |
J/hm2/a |
18199 |
3.14E+14 |
629.69 |
|
4 |
Ëŕ╦«ú║äŢ(shĘČ)─▄ |
6.49E+07 |
J/hm2/a |
27874 |
1.81E+12 |
3.62 |
|
5 |
Áěă˛Đşşh(huĘón) |
1.00E+10 |
J/hm2/a |
34377 |
6.79E+14 |
687.54 |
|
|
╣Ž─▄ú║͸Ϭşh(huĘón)ż│Ě■äŇ(wĘ┤)╣Ž─▄ |
|
|
|
|
7285.94 |
|
6 |
╦«ĚÍҢ░l(fĘí)(Ë├╦«┴┐) |
1.98E+10 |
J/hm2/a |
18199 |
3.60E+14 |
720.68 |
|
7 |
NPP |
1.19E+11 |
J/hm2/a |
13788 |
1.04E+15 |
3281.54 |
|
8 |
GPP |
6.31E+11 |
J/hm2/a |
2602 |
1.04E+15 |
3283.72 |
|
|
ŻY(jiĘŽ)śő(gĘ░u)ú║͸Ϭ╔˙ĹB(tĘĄi)┘Y«a(chĘún) |
|
|
|
|
10580.06 |
|
9 |
╗ţ╔˙╬´┴┐ |
1.69E+11 |
J/hm2 |
27576 |
4.16E+15 |
9320.69 |
|
10 |
▓ŢÁě╦«ĚÍ║Č┴┐ |
3.46E+10 |
J/hm2 |
18199 |
6.29E+14 |
1259.37 |
Îóú║1íó▒ż▒ÝĎď1hm2в▓ŢśËÁě×Ú└ř▀M(jĘČn)đđËő(jĘČ)╦Ń�����íú1hm2▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─˛î(qĘ▒)äË(dĘ░ng)┴Ž����íó╔˙ĹB(tĘĄi)Ě■äŇ(wĘ┤)╣Ž─▄╝░Ăń╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁÁ╚Ë┌Ş¸ÝŚ(xiĘĄng)─┐Á──▄ÍÁ│řĎďÍđç°(guĘ«)─▄ÍÁěŤÄ┼▒╚┬╩(5X1011sej/RMB,ç└(yĘón)├»│ČúČ2001)���ú╗
2�����íóNPP║═GPP╝░╗ţ╔˙╬´┴┐Á──▄ÍÁŮD(zhuĘún)ôQ┬╩üÝ(lĘói)Îď▒żĐđż┐�����úČĂń╦űÁ──▄ÍÁŮD(zhuĘún)ôQ┬╩üÝ(lĘói)ÎďË┌ç°(guĘ«)ŰH═ĘË├ÍŞś╦(biĘío)���ú╗
Ć─▒Ý1-33┐╔Ďď┐┤│÷�úČ1hm2▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)─࿨╠ß╣ęÁ─͸Ϭşh(huĘón)ż│Ě■äŇ(wĘ┤)╣Ž─▄╝s×Ú7285.94ď¬/hm2/a,����úČĂńÍđ╦«ĚÍҢ░l(fĘí)âr(jiĘĄ)ÍÁ×Ú720.68ď¬(em ¥)/hm2/a(┤╦Ě■äŇ(wĘ┤)╣Ž─▄┐╔Ďď╩╣şh(huĘón)ż│▒ú│ÍĎ╗ÂĘÁ─Ł˝Â╚úČŁMÎŃ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)äË(dĘ░ng)╬´║═Í▓╬´Á─đŔăˇ)�úČâ˘╔˙╬´┴┐(NPP)Á─âr(jiĘĄ)ÍÁ×Ú3281.54ď¬/hm2/a(┤╦ÝŚ(xiĘĄng)Ě■äŇ(wĘ┤)╣Ž─▄┐╔ĎďŁMÎŃ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)޸ţÉäË(dĘ░ng)╬´╝░╬ó╔˙╬´Á╚Á─đŔăˇ)úČ│§╝ë(jʬ)╔˙╬´╔˙«a(chĘún)┴Ž(GPP)Á─âr(jiĘĄ)ÍÁ×Ú3283.72ď¬(┤╦ÝŚ(xiĘĄng)Ě■äŇ(wĘ┤)╣Ž─▄┐╔ĎďŁMÎŃ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)▀\(yĘ┤n)ŮD(zhuĘún)���úČ╬Ř╩ŇCO2Á╚ÜÔˇw▓óˇwČF(xiĘĄn)Ď╗¤Á┴đ¤ÓŕP(guĘín)Á─╔˙ĹB(tĘĄi)Ě■äŇ(wĘ┤)╣Ž─▄)�íú
1hm2▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝s×Ú10580.06ď¬/hm2�����úČĂńÍđ╗ţ╔˙╬´┴┐Á─âr(jiĘĄ)ÍÁ╝s×Ú9320.69ď¬/hm2,░ÚŰSÍ°╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─▀M(jĘČn)╗»Â°đţĚeÁ─╦«ĚÍÁ─╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─âr(jiĘĄ)ÍÁ×Ú1259.37ď¬/hm2����íú
9.3.2 ňa┴Í╣¨└Ňв▓Ţ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁ
╬Ďéâ╝┘ďO(shĘĘ)ňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╗ÍĆ═(fĘ┤)Ă┌×Ú8─ŕúČ▓óô■(jĘ┤)┤╦ďu(pʬng)╣└▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁ���íú╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁV=Vs+Vc,╩ŻÍđVs▒Ý╩ż╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ě■äŇ(wĘ┤)╣Ž─▄Á─╠Š┤˙âr(jiĘĄ)ÍÁ����úČVc┤˙▒Ý╔˙ĹB(tĘĄi)┘Y▒żÁ─╠Š┤˙âr(jiĘĄ)ÍÁ����íú╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─Ě■äŇ(wĘ┤)╣Ž─▄Á─╠Š┤˙âr(jiĘĄ)ÍÁVs=1/2*T*(GPP*TrGPP /P+VNPP+V╦«)úČ╩ŻÍđGPP×Ú╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─│§╝ë(jʬ)╔˙«a(chĘún)┴Ž�����úČ°TrGPP ×ÚGPPÁ──▄ÍÁŮD(zhuĘún)ôQ┬╩, T×Ú╠Š┤˙Ľr(shʬ)Úg╗˛╗ÍĆ═(fĘ┤)Ľr(shʬ)Úg(replacement time), P×Ú─▄ÍÁ/ěŤÄ┼▒╚┬╩���úČVNPP║═V╦«ĚÍäe┤˙▒Ýâ˘╔˙╬´┴┐║═▓ŢÁě╦«ĚÍ┘A┴┐Á─âr(jiĘĄ)ÍÁ�����íúިô■(jĘ┤)╔¤╩÷ĚÍ╬÷����úČ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁV= 1/2*(GPP*TrGPP*T/P + T VNPP+ T V╦«)+VcúČËő(jĘČ)╦ŃŻY(jiĘŽ)╣űĎŐ▒Ý1-34�íú
▒Ý1-34 1hm2ňa┴Í╣¨└Ňв▓Ţ▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁďu(pʬng)╣└
|
ÝŚ(xiĘĄng) ─┐ |
Ëő(jĘČ)╦Ń╣ź╩Ż |
║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ(em$) |
éńÎó |
|
1.şh(huĘón)ż│Ě■äŇ(wĘ┤)╣Ž─▄╠Š┤˙âr(jiĘĄ)ÍÁ(Vs) |
Vs=1/2*T*(GPP*TrGPP /P+VNPP+Vˮ) |
29143.76 |
GPP║═TrGPPöÁ(shĘ┤)ô■(jĘ┤)üÝ(lĘói)ď┤Ë┌▒Ý5-2úČT×Ú8─ŕ�íú |
|
2.╔˙ĹB(tĘĄi)┘Y▒ż(Vc) |
Vc=Vc1+Vc2+íş+Vcn |
10580.06 |
Vc1,Vc2, íş,Vcn┤˙▒Ý▓╗═ČţÉđ═╔˙ĹB(tĘĄi)ŻMĚÍÁ─╔˙ĹB(tĘĄi)┘Y▒żíú |
|
┐é╠Š┤˙âr(jiĘĄ)ÍÁ |
V=Vs+Vc |
|
|
9.3.3 ňa┴Í╣¨└Ňв▓ŢśËÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁÁ─żC║¤╣└╦Ń
ď┌Ëő(jĘČ)╦Ń▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁĽr(shʬ)�����úČ╬Ďéâ╝┘ďO(shĘĘ)▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)═ŕ╚ź¤ű╩ž���íú╠Š┤˙âr(jiĘĄ)ÍÁ╩ă▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ě■äŇ(wĘ┤)âr(jiĘĄ)ÍÁ┼cÎď╚╗┘Y▒żâr(jiĘĄ)ÍÁÁ─┐é║═���íúިô■(jĘ┤)ňa┴Í╣¨└Ň├╦Á─îŹ(shʬ)ŰHúČ1hm2в▓ŢśËÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╠Š┤˙âr(jiĘĄ)ÍÁ╝s×Ú39724ď¬�ú╗Ć─24hm2śËÁěÁ─ăÚŤrüÝ(lĘói)┐┤úČĂń┐é╠Š┤˙âr(jiĘĄ)ÍÁ╝s×Ú953371ď¬(▒Ý1-35)�íú
▒Ý1-35 ňa┴Í╣¨└Ň24hm2┐╔└űË├▓Ţł÷(chĘúng)╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)âr(jiĘĄ)ÍÁżC║¤ďu(pʬng)╣└ŻY(jiĘŽ)╣ű
|
ÝŚ(xiĘĄng)─┐ |
├ŠĚe(hm2) |
║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ(ď¬) |
ć╬╬╗ |
|
Ď╗íóşh(huĘón)ż│Ě■äŇ(wĘ┤)╣Ž─▄ |
24 |
174862.56 |
em¥/a |
|
Â■����íó╔˙ĹB(tĘĄi)┘Y▒ż |
24 |
253921.44 |
em¥ |
|
╚říó┐é╠Š┤˙âr(jiĘĄ)ÍÁ |
24 |
953371.68 |
em¥ |
ިô■(jĘ┤)Ďď╔¤Áńđ═╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─Đđż┐ŻY(jiĘŽ)╣ű�úČ╬Ďéâ┐╔ĎďîŽ(duĘČ)ňa┴Í╣¨└Ň24hm2в▓ŢśËÁě▓Ţł÷(chĘúng)Á─âr(jiĘĄ)ÍÁθ│§▓Ż╣└╦Ńíú
9.4 đíŻY(jiĘŽ)
9.4.1 ─▄ÍÁ┼c▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ
▀\(yĘ┤n)Ë├─▄ÍÁ└ÝŇôîŽ(duĘČ)╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─âr(jiĘĄ)ÍÁ▀M(jĘČn)đđďu(pʬng)╣└���úČË├─▄ÍÁď¬(em$╗˛em¥)îŽ(duĘČ)╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─ŻY(jiĘŽ)śő(gĘ░u)╝░Ăńşh(huĘón)ż│Ě■äŇ(wĘ┤)╣Ž─▄Á─âr(jiĘĄ)ÍÁ▀M(jĘČn)đđďu(pʬng)╣└�����úČ┐╔Ďď╩╣âr(jiĘĄ)ÍÁďu(pʬng)╣└ŻĘ┴óď┌┐ĂîW(xuĘŽ)┐═Ë^Á─╗¨ÁA(chĘ│)Í«╔¤�íú▒żĐđż┐ŻY(jiĘŽ)╣ű▒Ý├¸�����úČď┌ňa┴Í╣¨└Ň���úČ1hm2▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ĂŻż¨├┐─ŕîŽ(duĘČ)ů^(qĘ▒)˲ŻŤ(jĘęng)Ł˙(jĘČ)켟I(xiĘĄn)Á─║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ╝s×Ú7285.94ď¬�����úČ°1hm2▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)͸Ϭ╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ╝s×Ú10580.06ď¬����íúĆ─24hm2śËÁěÁ─ăÚŤrüÝ(lĘói)┐┤�úČ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ě■äŇ(wĘ┤)╣Ž─▄Á─âr(jiĘĄ)ÍÁ╝s×Ú174863ď¬/aúČ╔˙ĹB(tĘĄi)┘Y▒żÁ─âr(jiĘĄ)ÍÁ╝s×Ú253941ď¬(▒Ý5-4)�íú╚š╣ű─│Ď╗ŻŤ(jĘęng)Ł˙(jĘČ)Ú_░l(fĘí)đđ×Úîž(dĘúo)Í┬▀@Ď╗ů^(qĘ▒)˲▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ôp╩žúČ╬Ďéâ┐╔ĎďË├╔¤╩÷Đđż┐ŻY(jiĘŽ)╣ű╣└╦Ńşh(huĘón)ż│Ě■äŇ(wĘ┤)╣Ž─▄╝░╔˙ĹB(tĘĄi)┘Y▒żÁ─ôp╩žăÚŤr�����úČĎ▓┐╔Ď直ô■(jĘ┤)╔˙╬´┴┐Á─ťp╔┘╝░GPPÁ─ÎâäË(dĘ░ng)ăÚŤrîŽ(duĘČ)╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─Ě■äŇ(wĘ┤)╣Ž─▄╝░╔˙ĹB(tĘĄi)┘Y▒żÎâäË(dĘ░ng)ăÚŤr▀M(jĘČn)đđďu(pʬng)╣└íú╚f(wĘĄn)Ď╗─│Ď╗Áěů^(qĘ▒)▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)═╦╗»���úČ╬Ďéâ┐╔Ď直ô■(jĘ┤)▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╣Ž─▄═╦╗»Á─│╠Â╚╣└╦Ń│÷ů^(qĘ▒)˲╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ôp╩žÁ─ăÚŤr�����íú
9.4.2 ─▄ÍÁ╔˙ĹB(tĘĄi)âr(jiĘĄ)ÍÁďu(pʬng)╣└ĚŻĚĘÁ─┐ĂîW(xuĘŽ)đď╝░Ŭ(yĘęng)Ë├ă░ż░
┼c鸯y(tĘ»ng)Á─ŻŤ(jĘęng)Ł˙(jĘČ)îW(xuĘŽ)ĚŻĚĘ║═ÍžŞÂĎÔďŞĚŻ╩Ż¤Ó▒╚�úČ▀\(yĘ┤n)Ë├─▄ÍÁ└ÝŇô▀M(jĘČn)đđ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─âr(jiĘĄ)ÍÁďu(pʬng)╣└Á─ĚŻĚĘŢ^×Ú┐═Ë^╣źŇř�����úČ┐╦Ě■┴╦鸯y(tĘ»ng)ĚŻĚĘÁ─͸Ë^╔ź▓╩����úČż▀Ëđç└(yĘón)├▄Á─┐ĂîW(xuĘŽ)╗¨ÁA(chĘ│)║═Ţ^×Ú┐═Ë^Á─âr(jiĘĄ)ÍÁ│▀Â╚úČϲ°ż▀ËđĆVĚ║Á─Ŭ(yĘęng)Ë├ă░ż░�����íú
9.5 ňa┴Í╣¨└Ň▓ŢÁěÍ▓▒╗îË╔˙╬´┴┐┘Y«a(chĘún)╝░ĂńĚ■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁäË(dĘ░ng)ĹB(tĘĄi)─úöM
9.5.1 ͸Ϭ╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░ÍěϬĚ■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁďu(pʬng)╣└ŻY(jiĘŽ)╣ű
═Ę▀^(guĘ░)îŽ(duĘČ)ňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ÄÎÝŚ(xiĘĄng)͸Ϭ╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─âr(jiĘĄ)ÍÁ▀M(jĘČn)đđďu(pʬng)╣└�����úČŻY(jiĘŽ)╣ű▒Ý├¸úĘ▒Ý1-36úęú║╔˙╬´┴┐╔˙ĹB(tĘĄi)┘Y«a(chĘún)(╗ţ╔˙╬´┴┐)Á─âr(jiĘĄ)ÍÁ×Ú1651â|ď¬╚╦├˝Ä┼úČ͸Ϭ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ě■äŇ(wĘ┤)╣Ž─▄Á─âr(jiĘĄ)ÍÁ└█Ëő(jĘČ)×Ú1651â|ď¬�����úČ͸Ϭ╠Š┤˙âr(jiĘĄ)ÍÁ8255â|ď¬�íú╠Š┤˙âr(jiĘĄ)ÍÁďu(pʬng)╣└ŻY(jiĘŽ)╣ű▒Ý├¸úČ╚š╣űňa┴ÍŞ˝└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)ďÔÁŻç└(yĘón)ÍěĂĂë─����úČϬ╗ÍĆ═(fĘ┤)╔¤╩÷ÄÎÝŚ(xiĘĄng)͸Ϭ╣Ž─▄Í┴╔┘đŔϬ═Â┘Y8255â|ď¬���íú
Ë╔Ë┌╩đł÷(chĘúng)╩žý`║═âr(jiĘĄ)Ş˝Ş▄ŚUÁ─┼Ąă˙�úČďSÂÓ╔˙ĹB(tĘĄi)ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ╬┤─▄ď┌鸯y(tĘ»ng)Á─ĚŻĚĘÍđÁ├ĎďˇwČF(xiĘĄn)����úČâr(jiĘĄ)ÍÁ▒╗Á═╣└┴╦úČâHÄÎÝŚ(xiĘĄng)͸ϬÁ─╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ě■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁúĘ1651â|ď¬/─ŕúęż═┤ˇ┤ˇ│Č▀^(guĘ░)┴╦ňa┴Í╣¨└ŇŮr(nĘ«ng)┴Í─┴ŁOśI(yĘĘ)«a(chĘún)ÍÁÁ─┐é║═úĘ33.97â|ď¬úę����íú×Ú┴╦ĆŤĐa(bĘ│)╩đł÷(chĘúng)ŻŤ(jĘęng)Ł˙(jĘČ)║═âr(jiĘĄ)Ş˝ˇw¤ÁÁ─╚▒¤ŢúČç°(guĘ«)╝ĎŬ(yĘęng)ÍĂÂĘ│÷Ď╗ÂĘÁ─Đa(bĘ│)âöÖC(jĘę)ÍĂ�����úČîŽ(duĘČ)▀@Ď╗ů^(qĘ▒)˲▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─ŻĘďO(shĘĘ)▒úÎo(hĘ┤)ËŔĎďÍž│Ííú
▒Ý1-36íí͸Ϭ╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░Ě■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁďu(pʬng)╣└▒Ý
|
âr(jiĘĄ)ÍÁţÉđ═ |
Ëő(jĘČ)╦Ń╣ź╩Ż |
âr(jiĘĄ)ÍÁúĘâ|ď¬úę |
|
1.╔˙╬´┴┐╔˙ĹB(tĘĄi)┘Y«a(chĘún) |
VQúŻ923.5QúĘď¬úę |
1651 |
|
2.Ě■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁ |
Es=Fc+Fo+So |
1651 |
|
2.1╣╠ÂĘCO2âr(jiĘĄ)ÍÁ |
Fc=1.63NPP*P=1.63*NPP*250(ď¬/t) =407.5NPP(ď¬) |
371 |
|
2.2ßîĚ┼O2âr(jiĘĄ)ÍÁ |
Fo=1.19NPP*Po=1.19*500NPP=595NPP(ď¬) |
541 |
|
2.3╠ß╣ęËđÖC(jĘę)╬´âr(jiĘĄ)ÍÁ |
SoúŻ923.5(J2+J4)úĘď¬úę |
739 |
|
3.╠Š┤˙âr(jiĘĄ)ÍÁ |
Vs= 1/2(Fc+Fo+So)*8+ VQ =4(Fc+Fo+So)+ VQ |
8255 |
9.5.2 ňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)╔˙╬´┴┐┘Y«a(chĘún)╝░͸ϬĚ■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁäË(dĘ░ng)ĹB(tĘĄi)Îâ╗»
ިô■(jĘ┤)╔˙╬´┴┐╝░͸Ϭ╣Ž─▄ÍÁÁ──úöMŻY(jiĘŽ)╣ű�����úČŻY(jiĘŽ)║¤╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░Ě■äŇ(wĘ┤)╣Ž─▄Ëő(jĘČ)╦ŃĚŻ│╠����úČ╬Ďéâ┐╔ĎďÁ├ÁŻ╔˙╬´┴┐╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░Ě■äŇ(wĘ┤)╣Ž─▄äË(dĘ░ng)ĹB(tĘĄi)Îâ╗»Á──úöMŻY(jiĘŽ)╣űúĘłD1-28úęíúĆ─łD

łD1-28 ňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)͸Ϭ╔˙ĹB(tĘĄi)╣Ž─▄ÍÁÎâ╗»äË(dĘ░ng)ĹB(tĘĄi)─úöMŻY(jiĘŽ)╣ű
Íđ┐╔Ďď┐┤│÷�����úČ1992─ŕďôů^(qĘ▒)▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Ë╔Ë┌Ţdđˇ┴┐▓ó╬┤─▄▀_(dĘó)ÁŻÎţ┤ˇÍÁ�����úČîŽ(duĘČ)▓ŢÁěÁ─╔˙╬´┴┐¤ű║─┴┐Á═Ë┌╔˙ÚL(zhĘúng)┴┐����úČ▓ŢÁěŢdđˇ┴┐╔đËđĎ╗ÂĘÁ─ď÷ÚL(zhĘúng)Łô┴Žíú▀@ĚNăÚŤr┼c1992─ŕĎďüÝ(lĘói)▓ŢđˇöÁ(shĘ┤)┴┐Á─ď÷ÚL(zhĘúng)Á─╩┬îŹ(shʬ)╦¨ÎC├¸�����úČŇf(shuĘş)├¸┤╦─úöMŻY(jiĘŽ)╣ű┼c╩┬îŹ(shʬ)¤ÓĚű�����íú
─úöMŻY(jiĘŽ)╣ű▒Ý├¸úČĆ─┐éˇwăÚŤr┐┤���úČÎďĆ─1992─ŕĎďüÝ(lĘói)���úČňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Í▓▒╗îË╔˙╬´┴┐╔˙ĹB(tĘĄi)┘Y«a(chĘún)╝░͸ϬĚ■äŇ(wĘ┤)╣Ž─▄Á─âr(jiĘĄ)ÍÁ│╩ČF(xiĘĄn)│÷ď÷ÚL(zhĘúng)ĹB(tĘĄi)äŢ(shĘČ)úČÁŻ2004─ŕ�úČŞ¸ÝŚ(xiĘĄng)Ě■äŇ(wĘ┤)╣Ž─▄ÍÁ▓ó╬┤▀_(dĘó)ÁŻÝöśOáţĹB(tĘĄi)úČď÷ÚL(zhĘúng)╔đËđĎ╗ÂĘÁ─Łô┴Ž���íú2004─ŕúČ╔˙╬´┴┐╔˙ĹB(tĘĄi)┘Y«a(chĘún)Á─âr(jiĘĄ)ÍÁ×Ú2421â|ď¬���úČ͸ϬĚ■äŇ(wĘ┤)╣Ž─▄âr(jiĘĄ)ÍÁ1902â|ď¬/─ŕúĘĂńÍđ����úČ╣╠ÂĘCO2âr(jiĘĄ)ÍÁ407â|ď¬�úČßîĚ┼O2âr(jiĘĄ)ÍÁ595â|ď¬úČ╠ß╣ęËđÖC(jĘę)╬´âr(jiĘĄ)ÍÁ900â|ď¬úę���úČ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁ▀_(dĘó)ÁŻ10031â|ď¬���íúިô■(jĘ┤)─úöMţA(yĘ┤)ťy(cĘĘ)�úČ2031─ŕ�úČňa┴Í╣¨└Ň▓ŢÁě╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─͸Ϭ╔˙ĹB(tĘĄi)ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁîó▀_(dĘó)ÁŻú║╔˙ĹB(tĘĄi)┘Y«a(chĘún)(╔˙╬´┴┐)Á─║ŕË^ŻŤ(jĘęng)Ł˙(jĘČ)âr(jiĘĄ)ÍÁ×Ú2585â|ď¬úČ͸ϬĚ■äŇ(wĘ┤)╣Ž─▄Á─âr(jiĘĄ)ÍÁ×Ú1928â|ď¬/a����úČ─ăĽr(shʬ)úČ╔˙ĹB(tĘĄi)¤ÁŻy(tĘ»ng)Á─╠Š┤˙âr(jiĘĄ)ÍÁ×Ú10296â|ď¬�íú
╔¤Ď╗Ýô(yĘĘ) [1] [2] [3] [4] [5] [6]
|